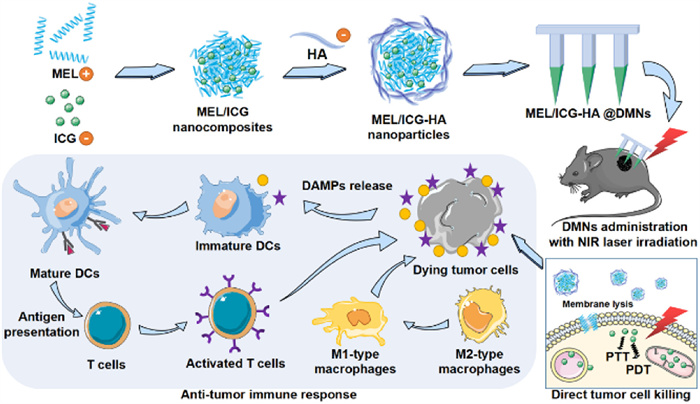
Scheme 1.
Schematic illustration of DMNs-assisted in situ cancer vaccine combined with cytolytic peptide for antitumor immunotherapy.
Dissolving microneedle-assisted in situ cancer vaccine combined with cytolytic peptide for anti-melanoma immunotherapy
Ying Sun , Minglong Chen , Ying Chen , Wanchen Zhao , Yanping Fu , Zhengwei Huang , Chao Lu , Chuanbin Wu , Xin Pan , Guilan Quan

Immunotherapy has emerged as a pivotal approach in cancer treatment, primarily by activating and enhancing the immune system to specifically target and eliminate tumor cells [1]. Several immunotherapeutic strategies, including checkpoint inhibitors, chimeric antigen receptor (CAR) T-cell therapy, cytokine therapy, and cancer vaccines, have been approved and have demonstrated promising clinical outcomes [2–6]. Among these, cancer vaccines hold particular promise due to their ability to induce cancer-specific immune responses and establish long-term immune memory, thereby helping to prevent tumor recurrence [7]. However, the clinical efficacy of cancer vaccines has been limited by challenges such as weak immunogenicity, patient tumor heterogeneity, the immunosuppressive tumor microenvironment (TME), and obstacles in vaccine delivery [2,8–10].
For a cancer vaccine to elicit an effective antitumor response, it must successfully promote the release and presentation of highly immunogenic antigen. Recent research has shown that immunogenic cell death (ICD) not only induce tumor cell death but also promote the release of tumor neoantigens and damage-associated molecular patterns (DAMPs), thereby significantly enhancing the immune system's ability to recognize and eliminate tumors [11,12]. During ICD, dying tumor cells release DAMPs, including calreticulin (CRT) and heat shock proteins (HSPs), which bind to pattern recognition receptors on antigen-presenting cells (APCs). This interaction triggers a cascade of cellular processes that activate both innate and adaptive immune responses, thereby addressing the challenge of weak immunogenicity in cancer vaccines [13–15]. Photothermal therapy (PTT) offers a promising strategy for antitumor therapy, which utilizes photosensitizers to convert near-infrared (NIR) light energy into hyperthermia [16,17]. PTT can directly ablate cancer cells while also inducing the ICD effect, thus providing a rich source of tumor-associated antigens that serve as an in situ whole-cell vaccine [18,19]. This approach has the potential to generate a robust and specific antitumor immune response, addressing the challenges of tumor heterogeneity and enhancing the overall efficacy of cancer vaccines.
Despite the potential of cancer vaccines to induce tumor-specific immune responses, their clinical efficacy has been hampered by the immunosuppressive TME. Key components of the TME, such as regulatory T cells, myeloid-derived suppressor cells, and tumor-associated macrophages (TAMs), inhibit effective antitumor responses [20]. TAMs, in particular, play a critical role in tumor immune evasion. While M1-TAMs exhibit tumor-killing properties with strong antigen-presenting capacity and immune-promoting antitumor activity, M2-TAMs support tumor growth and metastasis by secreting proangiogenic factors [21,22]. Thus, the ability to regulate TAM polarization from the M2 to the M1 phenotype can significantly improve immunosuppressive TME, thereby enhancing the efficacy of cancer vaccines. Melittin (MEL), the primary component of honeybee venom, has shown potent antitumor activity by disrupting biological membranes. In this case, MEL would act as a membranolytic weapon to potentiate the PTT by increasing the cellular uptake of photosensitizer [23,24]. Notably, MEL has been demonstrated to exert significant immunomodulatory effects, which can enhance antitumor immunity [25,26]. Recent studies have demonstrated that MEL can selectively target M2-TAMs and induce their polarization to the M1 phenotype, thereby improving the tumor-suppressive immune microenvironment and enhancing CD8+ T cell-mediated tumor killing [27,28]. Furthermore, MEL has been shown to induce the formation of tumor cell vesicles that can activate APCs in lymph nodes, thus inducing a systemic antitumor immune response [29,30]. Thus, incorporating strategies like MEL-mediated TME modulation was expected to address the current limitations of cancer vaccines and improve their clinical outcomes.
Targeted delivery of ICD inducers to the TME is essential for in situ generation of robust cancer vaccines. Dissolving microneedles (DMNs) offer a promising solution by delivering ICD inducers directly to the superficial tumor site in a targeted, non-invasive manner [31,32]. Consisting of hundreds of micro-sized needle tips neatly arranged on a substrate, DMNs can penetrate the skin's stratum corneum barrier, deliver drugs via microchannels precisely to the superficial lesion, and leverage the immune-rich environment of the epidermis and dermis to enhance antitumor responses. This approach may promote localized tumor antigen release, improving vaccination efficacy while minimizing systemic toxicity [33,34].
Herein, based on the PTT-induced in situ cancer vaccine strategy and MEL-enhanced antitumor immune responses, we developed a nanocomplex-loaded DMN delivery system for treating superficial melanoma (Scheme 1). Specifically, positively charged MEL was integrated with the negatively charged photosensitizer indocyanine green (ICG) to form nanocomposites (MEL/ICG) through electrostatic interaction. These nanocomposites were then wrapped by hyaluronic acid (HA) to formulate nanocomplexes (MEL/ICG-HA), which were further encapsulated into DMN patches (MEL/ICG-HA@DMNs). This system aims to achieve: (1) Direct tumor ablation through PTT-induced hyperthermia assisted by MEL-generated membrane lysis. (2) Activation of antitumor immune response via PTT-induced ICD effect to form in situ whole-cell vaccine. (3) Enhanced antitumor immunity through MEL-induced immunomodulation to prevent tumor recurrence. Our results showed that MEL/ICG-HA@DMNs could effectively concentrate the drug at the tumor site, raise the local temperature, and successfully ablate the tumor. Furthermore, MEL/ICG-HA@DMNs promoted the maturation of dendritic cells (DCs) and increased the proportion of M1/M2-type macrophages in draining lymph nodes, indicating a robust antitumor immune response. This system addressed the limitations of traditional cancer vaccines by ensuring precise, targeted delivery and robust immune activation. Moreover, due to the unique cytotoxic effect of MEL and PTT, this system may also have applications in treating local infections, highlighting its promising clinical potential.
The MEL/ICG-HA was prepared through a self-assembly process facilitated by electrostatic interactions. MEL, a highly positively charged polycation, was observed to form MEL/ICG nanocomplexes with negatively charged ICG. Subsequently, HA was employed to wrap around the MEL/ICG complex, thereby enhancing its stability (Fig. 1A). The transmission electron microscope (TEM) examination revealed that the MEL/ICG-HA exhibited a uniform particle size and regular spherical morphology (Fig. 1B). The average diameter of MEL/ICG-HA was approximately 116.80 ± 1.19 nm with a polymer dispersity index (PDI) of 0.16 ± 0.01 (Fig. 1C), which was slightly larger than that of MEL/ICG (94.68 ± 2.45 nm) (Fig. S1 in Supporting information). The zeta potential of MEL/ICG-HA was approximately −26.7 ± 2.45 mV (Fig. S1). The encapsulation efficiency of MEL and ICG was 15.98% ± 3.57% and 24.96% ± 6.92%, respectively. Following storage at 4 ℃ for 7 days in the absence of light, the particle size of MEL/ICG-HA showed minimal change and remained homogeneous, indicating favorable storage stability (Fig. S2 in Supporting information). This result suggests that the formulation could be suitable for further experiments.
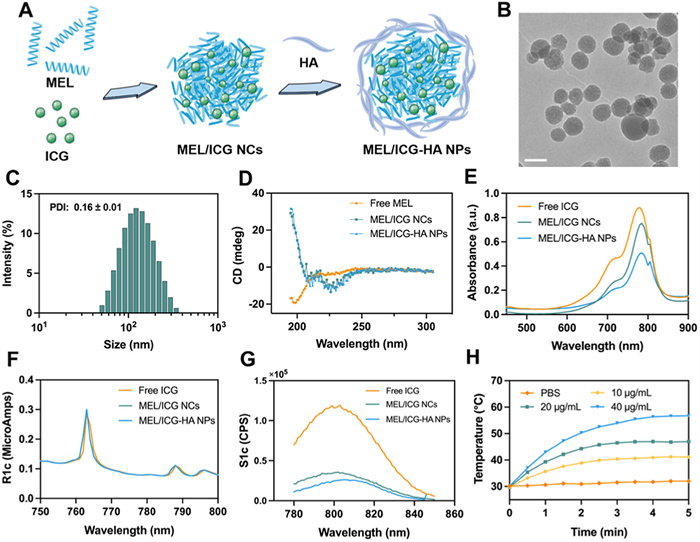
As illustrated in the circular dichroism (CD) spectra (Fig. 1D), free MEL exhibits a pronounced negative signal at 200 nm, indicative of its random curling configuration within the PBS solution. In contrast, MEL/ICG and MEL/ICG-HA showed two negative shoulder peaks at 208 and 222 nm, which is the characteristic peak pattern of the α-helix structure. These findings demonstrate that the complexation of MEL and ICG can alter the conformation of MEL. Compared with free ICG, both MEL/ICG and MEL/ICG-HA exhibited similar characteristic absorption peaks, thereby indicating the successful complexation of ICG with MEL (Fig. 1E). Moreover, the fluorescence properties of ICG remained unaltered following the formation of the complex, as evidenced by the observation that MEL/ICG-HA exhibited a prominent absorption peak at 763 nm in the excitation spectrum and 805 nm in the emission spectrum, which aligns with the characteristics of free ICG (Figs. 1F and G). The Fourier transform infrared spectroscopy (FTIR) characteristic peaks of MEL, ICG, and HA were discernible in MEL/ICG-HA (Fig. S3 in Supporting information), further substantiating the successful preparation of MEL/ICG-HA.
The hemolysis assay was performed to ascertain whether MEL/ICG and MEL/ICG-HA exerted any deleterious effects on RBC integrity. The free MEL exhibited a pronounced hemolytic effect. The hemolysis rate was 9.74% for MEL/ICG and 4.98% for MEL/ICG-HA at a concentration of 25 µg/mL, respectively (Fig. S4 in Supporting information). The observed reduction in hemolysis may be attributed to the complexation of ICG and HA, which has the effect of decreasing the affinity of MEL for erythrocytes. To assess the photothermal properties of ICG, a MEL/ICG-HA solution was diluted into varying concentrations and subjected to laser irradiation (808 nm, 1 W/cm2) for 5 min. As illustrated in Fig. 1H, the temperature of MEL/ICG-HA increased rapidly to 45 ℃ and remained above the damage threshold of tumor cells (42 ℃). This increase in temperature was found to be concentration-dependent, which was consistent with the visual thermal image of the MEL/ICG-HA solution (Fig. S5 in Supporting information). These findings substantiate the assertion that MEL/ICG-HA exhibits remarkable photothermal properties.
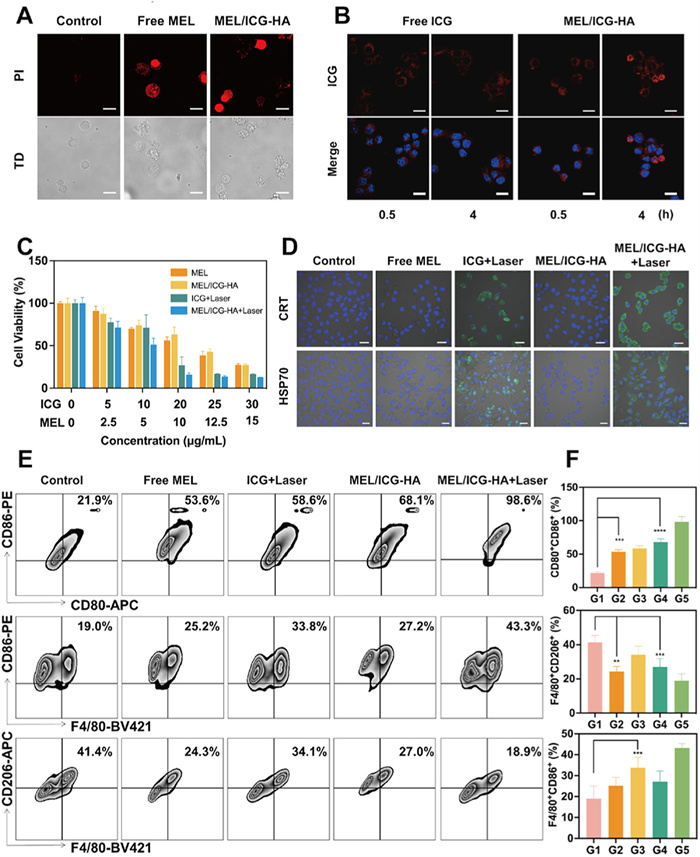
Encouraged by the excellent photothermal conversion performance, an investigation was conducted to examine the cellular accumulation behavior of MEL/ICG-HA in B16 cells. To ascertain whether MEL/ICG-HA possesses cytolytic ability, B16 cells were co-cultured with MEL/ICG-HA, and the membrane integrity was observed via confocal laser scanning microscope (CLSM). As illustrated in Fig. 2A, a pronounced PI fluorescence signal was observed in the B16 cells that were in contact with MEL. Furthermore, a notable fluorescence signal was exhibited in the cells that had been co-incubated with MEL/ICG-HA, which was accompanied by membrane lysis and foaming. These results demonstrate that MEL/ICG-HA has the capacity to remarkedly disrupt the membrane structure of tumor cells. Compared with free ICG, MEL/ICG-HA demonstrated a greater accumulation in B16 cells, exhibiting a time-dependent enhancement in fluorescence intensity within 4 h (Fig. 2B). This result may be attributed to the enhanced membrane permeation and subsequent increased intracellular uptake of ICG resulting from the cytolytic effect of MEL. To further assess the intracellular distribution of MEL/ICG-HA, the cell nucleus, lysosomes, and mitochondria were labeled by fluorescent staining. The green fluorescence signals of ICG appeared uniformly in the cytoplasm, partially overlapping with the red fluorescence signal of labeled lysosomes and highly overlapping with labeled mitochondria (Fig. S6 in Supporting information), indicating that MEL/ICG-HA were primarily distributed in the mitochondria within B16 cells.
ICG exhibits a pronounced absorption peak at 785 nm, which can facilitate the generation of hyperthermia and reactive oxygen species (ROS), thereby imparting a dual cell-killing effect that combines PTT with photodynamic therapy [35,36]. Herein, DCFH-DA, a fluorescent probe that is sensitive to oxidation, was employed to detect the ROS generation in B16 cells. As illustrated in Fig. S7 (Supporting information), following NIR laser irradiation, B16 cells treated with free ICG and MEL/ICG-HA displayed a distinct green fluorescence signal, and the fluorescence intensity in cells of the MEL/ICG-HA group was greater than that of the ICG group. These results indicated that MEL/ICG-HA could generate ROS in cells via laser irradiation, thereby inducing oxidative stress-mediated cell death.
As illustrated in Fig. S8A (Supporting information), in the absence of laser irradiation, the cell viability of B16 cells treated with free ICG was found to be 85% at concentrations up to 40 µg/mL, indicating the absence of dark cytotoxicity. In contrast, when laser irradiation was applied, a significant decrease in viability rate was observed with increasing concentration (Fig. 2C). This phenomenon may be attributed to the concentration-dependent photothermal conversion property. B16 cells treated with low concentrations of MEL exhibited no apparent cell death, while the cell viability rate decreased to 16% at concentrations exceeding 20 µg/mL (Fig. S8B in Supporting information). However, there was no notable change in cell viability as the concentration increased further. This result may be associated with the mechanism of membrane damage induced by MEL [37]. At low concentrations, MEL remains parallel to the cell membrane surface without exhibiting cytolysis toxicity. At a certain concentration, the hydrophobic fragments of MEL insert into the phospholipid bilayer, forming multimers that perforate the membrane and induce cell apoptosis or necrosis. Following NIR irradiation, B16 cells treated with MEL/ICG-HA exhibited a superior cell killing effect compared to MEL or phototherapy alone, with a cell viability of only 15.9% at an ICG concentration of 20 µg/mL. This result may be attributed to the enhanced ICG cellular amount resulting from the membrane-breaking action of MEL, which was consistent with the cellular uptake result.
Previous studies have demonstrated that phototherapy can induce damage to tumor cells through the generation of hyperthermia and ROS, as well as the stimulation of the ICD effect, thereby activating the antitumor immune response. During ICD, dying tumor cells release a series of danger signals [14,38]. To ascertain whether phototherapy could trigger the ICD effect, specific ICD markers, including CRT exposure and HSP70 expression, were analyzed. Following treatment with ICG and MEL/ICG-HA in conjunction with laser irradiation, a pronounced fluorescent signal of CRT was observed on the surface of the cell membrane, indicating that MEL/ICG-HA induced CRT translocation under laser irradiation. Furthermore, HSP70 expression in B16 cells was significantly upregulated. These results substantiate that MEL/ICG-HA can induce the ICD effect in tumor cells via phototherapy (Fig. 2D).
These DAMPs, including CRT and HSP70, act as “eat me” signals to recruit APCs and promote antigen presentation [39]. Flow cytometry analysis was employed to assess whether ICD induced by MEL/ICG-HA-mediated phototherapy could generate in situ cancer vaccine to activate the immune response. The signature proteins of mature DCs, CD80 and CD86, were detected. The percentage of double-positive cells was 52.7% in the group where B16 cells were pre-treated with free ICG plus laser irradiation, followed by coincubation with DCs (Figs. 2E and F). This result indicates that PTT-induced ICD in tumor cells can facilitate DC maturation, potentially due to the release of tumor antigens. Additionally, B16 cells pre-treated with MEL also demonstrated enhanced DC maturation, which is likely attributable to membrane damage caused by MEL, leading to the release of intracellular tumor-associated antigens. B16 cells pre-treated with MEL/ICG-HA plus laser irradiation exhibited a significant increase in CD80+CD86+ DCs, with a rate of 98.4%, indicating successful immune activation induced by phototherapy-mediated in situ cancer vaccination.
In addition, macrophages are integral to the ICD-induced antitumor immune response. M1-type macrophages are capable of phagocytosing dead or dying tumor cells and presenting a substantial quantity of antigenic information, which promotes the DC maturation. In contrast, M2-type macrophages have been demonstrated to suppress antitumor immunity [40]. As illustrated in Figs. 2E and F, in the co-incubation system, all treatment groups upregulated the proportion of M1-type macrophages (F4/80+CD86+) and decreased the proportion of M2-type macrophages (F4/80+CD206+) in comparison to the untreated group. Specifically, the ICG + laser (G3) and MEL/ICG-HA+ laser (G5) treatments resulted in an increase in the proportion of F4/80+CD86+ double-positive macrophages in bone marrow-derived macrophages (BMDMs), reaching 33.8% and 43.3%, respectively. While, the proportion of F4/80+CD206+ macrophages in BMDMs following ICG + laser (G3) and MEL/ICG-HA + laser (G5) treatments was 34.1% and 18.9%, respectively, which was significantly lower than that observed in the untreated group. These results suggest that PTT can effectively induce ICD in tumor cells, thereby stimulating macrophage activation and differentiation into M1-type macrophages. It is noteworthy that free MEL treatment also promoted the polarization of M2-type macrophages toward the M1-type, thereby contributing to the disruption of the immunosuppressive microenvironment and enhancing the responsiveness of the antitumor immune response.
The MEL/ICG-HA@DMNs were fabricated by a multi-step centrifugation molding technology as previously reported [41]. As illustrated in Figs. 3A and B, the fabricated MEL/ICG-HA@DMNs arrays exhibited a well-defined quadrangular cone and prismatic splicing structure, with the needles exhibiting a sharp, defined morphology. The loaded drug was concentrated at the tip of the needles, as evidenced by the 3 dimensions (3D) reconstruction images obtained via CLSM (Figs. 3C and D). The ability to puncture the skin stratum corneum with sufficient mechanical strength is a prerequisite for transdermal drug delivery. The MEL/ICG-HA@DMNs patch was observed to create a skin puncture rate of up to approximately 98%. Additionally, the hematoxylin & eosin (H&E) staining image of skin sections revealed the presence of obvious pores after DMNs insertion (Fig. S9 in Supporting information). These results indicate an efficient skin puncture ability for drug delivery.
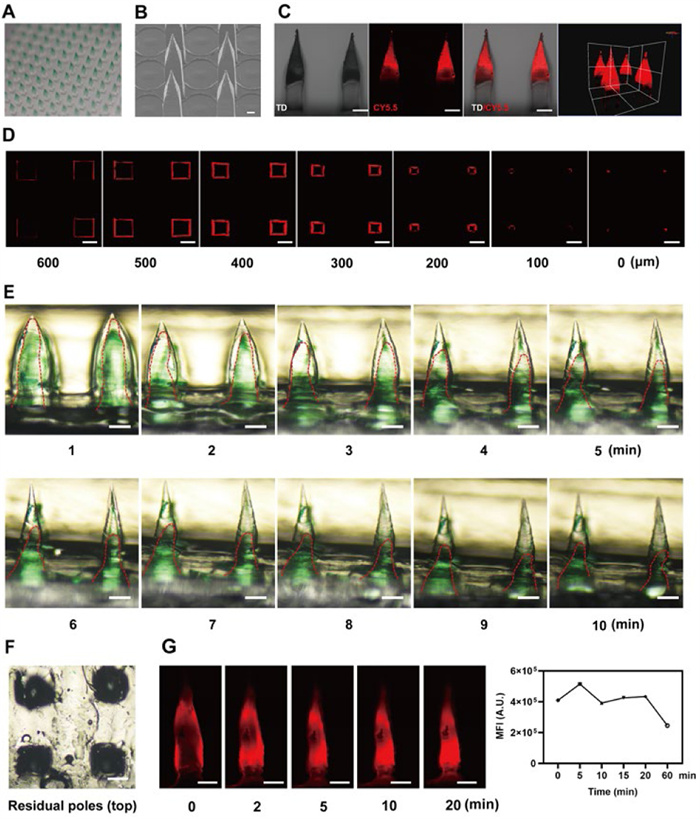
To investigate the dissolving property, the MEL/ICG-HA@DMNs patch was pressed into the rat skin for 2 min, after which the substrate was removed for observation. The negligible remaining portion of the needle body on the base indicated the complete dissolving of MEL/ICG-HA@DMNs, which is beneficial for precise and efficient drug delivery (Fig. S10 in Supporting information). Furthermore, a gelatin skin model with comparable water content and elasticity to human skin was constructed to examine the dissolving process of MEL/ICG-HA@DMNs. As illustrated in Figs. 3E and F, the height of the needle tips diminished rapidly within 5 min after insertion into the gelatin model, and only a portion of the needle body remained at 10 min. As shown in Fig. S11 (Supporting information), the particle size of MEL/ICG-HA NPs exhibited a slight increase upon the dissolution of MEL/ICG-HA@DMNs, which could be attributed to the additional encapsulation of HA during the preparation process of DMNs. These results indicated that MEL/ICG-HA@DMNs exhibited excellent dissolving properties, suggesting its potential for efficient drug delivery.
The long-term accumulation of photosensitizers has been demonstrated to enhance tumor-killing efficacy. As illustrated in Fig. 3G, the fluorescence intensity increased with the prolongation of the insertion time, indicating that the loaded drug was released from the DMNs concomitant with the dissolving. After insertion for 20 min, the residual microneedle base was removed, yet the fluorescent signal remained observable in the gelatin model at 60 min. This result implied that the drug released from the microneedle could be retained at the administration site for an extended period of time. Furthermore, to investigate the diffusion and permeation of the drug from MEL/ICG-HA@DMNs in the skin, the inserted site of isolated rat skin was imaged via CLSM. Following the insertion of DMNs, the formation of visible pores at the puncture site was observed, accompanied by the presence of fluorescent signals around the microchannels. With the prolongation of time, the fluorescent signals were observed to penetrate deeper into the skin (Fig. 4A). The layer-by-layer scanning and 3D construction images exhibited that the fluorescence signal could be observed beyond the height of the microneedle (1200 µm) 10 min post-application (Fig. 4B), indicating that MEL/ICG-HA@DMNs were capable of delivering loaded drugs to the deeper layers of the skin. These results demonstrate that MEL/ICG-HA@DMNs possess advantageous properties, including rapid dissolving, prolonged accumulation, and deep penetration, which collectively facilitate efficient transdermal drug delivery.
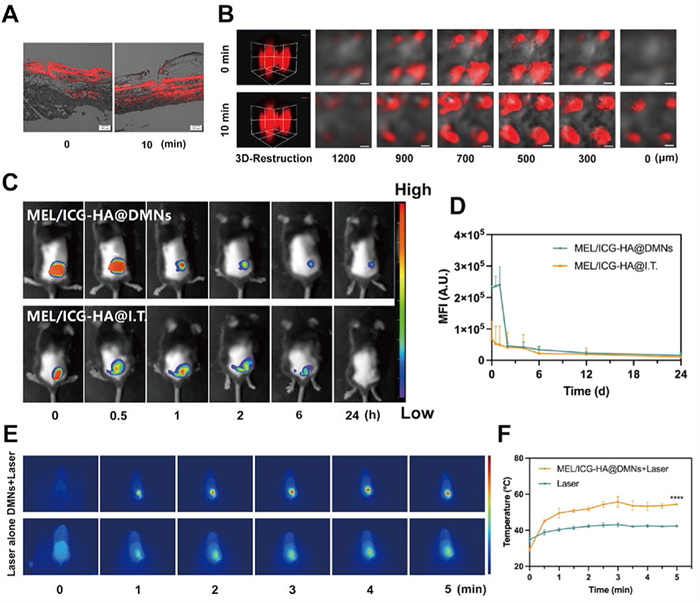
Considering the natural targeting effect of DMNs for superficial tumors, we further investigated the topical accumulation and biodistribution of MEL/ICG-HA@DMNs in melanoma-bearing mice using a live fluorescence imaging system. As illustrated in Figs. 4C and D, a pronounced fluorescence signal was observed at the tumor site following administration of MEL/ICG-HA@DMNs. In contrast, while fluorescent signals remained visible 24 h post-administration with DMNs, no such fluorescent signals were detected in mice receiving intra-tumoral injections. The results indicated that microneedle transdermal delivery can facilitate targeted and concentrated drug delivery to the tumor site, prolonging its retention at the site and enhancing the efficacy of PTT while minimizing damage to normal tissues.
The in vivo photothermal conversion capacity of MEL/ICG-HA@DMNs was subsequently investigated in melanoma-bearing mice. Animal experiments were permitted by the Institute Animal Care and Use Committee of Sun Yat-sen University. Previous studies have demonstrated that a treatment temperature exceeding 42 ℃ is required to induce apoptosis in tumor cells, while a temperature above 45 ℃ is sufficient to ablate the tumor [16]. The DMN patch was then directly inserted into the tumor site, followed by irradiation for 5 min (808 nm, 1 W/cm2). As shown in Figs. 4E and F, the local temperature at the tumor site increased slightly following laser irradiation alone, which remained below the threshold for thermally induced cellular damage. In contrast, the local temperature of the tumor site after MEL/ICG-HA@DMNs administration was rapidly increased to 45 ℃ within 30 s under NIR irradiation, which was higher than the damage threshold of the tumor, and remained above 48 ℃ for 5 min after continuous irradiation. Once the laser was turned off, the temperature quickly declined to the surrounding temperature within a few seconds. Consequently, MEL/ICG-HA@DMNs were able to successfully deliver nanoparticles to the local tumor with a notable in vivo photothermal conversion efficiency, which can effectively exert PTT anti-tumor effects.
Subsequently, the antitumor efficacy of MEL/ICG-HA@DMNs was investigated in a melanoma-bearing mouse model (Fig. 5A). The mice were randomly assigned to six groups. G1 was the control group and received no treatment. The treatment groups were as follows: G2: MEL@DMNs, G3: MEL/ICG-HA@DMNs, G4: ICG@DMNs + laser, G5: MEL/ICG-HA@DMNs + laser, and G6: MEL/ICG-HA@I.T. + laser. All treatment groups exhibited anti-tumor efficacy in comparison to the control group. As illustrated in Fig. 5B, the mice in the MEL@DMNs and MEL/ICG-HA@DMNs groups exhibited a slow increase in tumor growth up to day 6 post-administration, followed by an accelerated growth in the later stages. This indicated that the antitumor efficacy of a single MEL treatment was constrained. In contrast, the ICG@DMNs + laser, MEL/ICG-HA@DMNs + laser, and MEL/ICG-HA@I.T. + laser groups effectively regressed the tumor growth, indicating that PTT can exert strong antitumor efficacy. Notably, the MEL/ICG-HA@DMNs + laser group exerted distinct tumor eradication with nearly no relapse throughout the treatment period. This finding indicated that the combination of MEL and PTT can enhance the antitumor efficacy, and that the microneedle transdermal delivery system can precisely and uniformly deliver the drug to the tumor lesion, thereby optimizing the effectiveness of PTT.
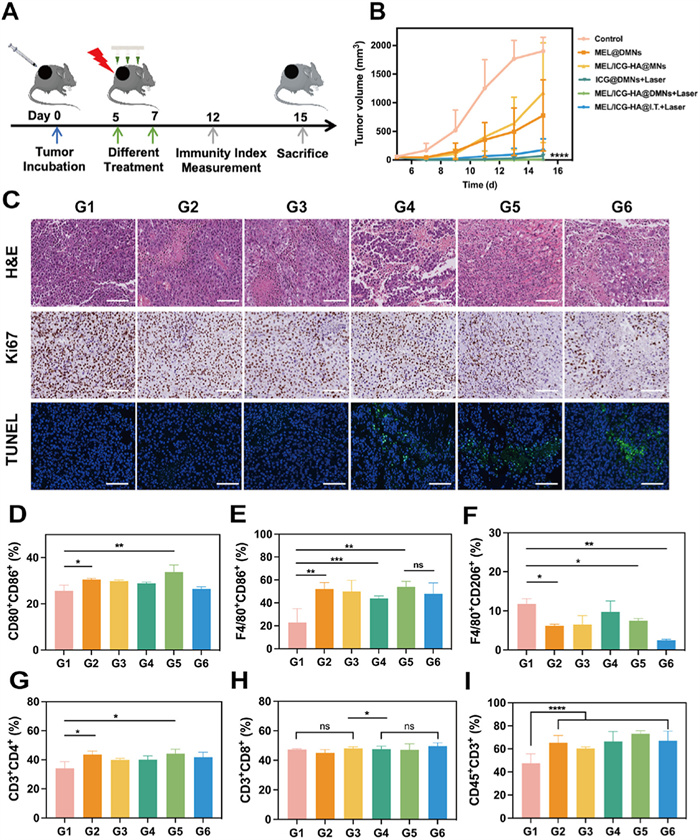
Histological analysis of the tumor tissues was conducted using H&E, Ki67, and TdT-mediated dUTP nick-end labeling (TUNEL) staining. As depicted in Fig. 5C, H&E staining revealed that the tumor cells in the G1 group were tightly arranged with well-preserved morphology. In contrast, the tumor cells in the G2 and G3 groups exhibited irregular edges and incomplete cytoplasmic staining, suggesting potential membrane damage and leakage of intracellular contents. The G4–G6 groups showed a loose cell arrangement, cellular crumpling, and a reduced nucleus-to-cytoplasm ratio, indicating further damage to the tumor tissue. Ki67 staining demonstrated a significant reduction in the positive indexes across the treatment groups, with the most notable decrease observed in the G5 and G6 groups. This finding suggests that the combined treatment of MEL and PTT was the most effective in inhibiting tumor cell proliferation. TUNEL staining revealed the presence of green fluorescent signals in the G4–G6 groups, indicative of the effective induction of apoptosis. Collectively, these results indicated that the combination of MEL/ICG-HA@DMNs and laser irradiation exhibited superior antitumor effects by inhibiting tumor cell proliferation and inducing apoptosis.
To exploit the mechanisms underlying the satisfying antitumor efficacy, we conducted a flow cytometric analysis of single-cell suspensions derived from the draining lymph nodes of mice subjected to different treatments. The analysis included the phenotypes and population of DCs, macrophages, and T cells. As shown in Fig. 5D and Fig. S12A (Supporting information), the proportion of CD80+CD86+ DCs increased to 33.8% following MEL/ICG-HA@DMNs + Laser treatment. Meanwhile, it was found that the MEL@DMNs treatment resulted in a notable elevation in the proportion of CD80+CD86+DCs, which could be attributed to the stimulatory effect of MEL as a host defense peptide on the immune system. As illustrated in Figs. 5E and F and Fig. S12B (Supporting information), the population of M1-type macrophages (F4/80+CD86+) was significantly elevated in all the treatment groups in comparison to the untreated group. Additionally, the MEL-treated group exhibited a notable reduction in the proportion of M2-type macrophages. These results indicate that MEL and PTT can promote tumor antigen release during tumor cell destruction, which in turn stimulates the proliferation and differentiation of macrophages into the M1 type, enhancing macrophage phagocytosis and antigen presentation. Furthermore, as shown in Figs. 5G–I and Figs. S12C and D (Supporting information), CD45+CD3+ T cells were significantly more prevalent in the lymph nodes of treated mice compared to the untreated group, with the most pronounced increase observed in the MEL/ICG-HA@DMNs + laser group. These results indicate that the combined application of MEL and PTT can facilitate the maturation of activated DCs, promote the conversion of M2-type macrophages to M1-type macrophages, regulate the inhibitory effects of M2-type macrophages, and activate T cells, thereby eliciting specific antitumor immune responses.
In order to facilitate the clinical translation of this technology, the biosafety of the MEL/ICG-HA@DMNs was investigated. Firstly, the process of skin recovery was visualized following the application of DMNs to the dorsal skin of healthy C57BL/6 mice. As illustrated in Fig. S13A (Supporting information), the MEL/ICG-HA@DMNs patch itself induced only minor trauma to the skin at the administration site, and the resulting damage to the skin barrier was transient and reversible. Although the PTT treatment following MEL/ICG-HA@DMNs application resulted in a certain degree of skin burn and disruption of the skin barrier, the skin was able to return to normal relatively quickly after administration.
The body weight of the mice was monitored throughout the course of the antitumor experiments. As illustrated in Fig. S13B (Supporting information), the mice in the control group exhibited weight gain on day 6, which is likely due to the accelerated growth of the tumor. Subsequently, they experienced a weight loss, which could be attributed to the tumor's excessive size at the later stage, thereby impacting the mice's normal life activities. With the exception of the untreated control group, no statistically significant change in body weight was observed in the other treatment groups. The histological analysis of the major organs (liver, kidney, spleen, lung, and heart) demonstrated that no discernible damage or abnormalities were observed in the mice treated with MEL/ICG-HA@DMNs with or without NIR irradiation (Fig. S13C in Supporting information). Furthermore, the blood biomarkers of mice treated with MEL/ICG-HA@DMNs +Laser, including the red blood cell count, white blood cell count, platelet count, as well as the liver and kidney function index, were all within the corresponding normal ranges (Fig. S14 in Supporting information), indicating that the treatment did not induce any adverse effects on the mice. These findings collectively substantiate the excellent biocompatibility of the MEL/ICG-HA@DMNs system.
In this study, we developed rapidly-dissolving DMNs that integrate the positively charged cytolytic peptide MEL and the negatively charged photosensitizer ICG via a facile and controllable fabrication method. The excellent antitumor efficacy of the constructed system could be attributed to a combined effect of PTT, a membranolytic effect, and immunotherapy. (ⅰ) Direct tumor ablation via hyperthermia generated by ICG under NIR irradiation. (ⅱ) The membranolytic effect assisted by MEL was capable of disrupting cell membranes and thereby enhancing the efficacy of PTT. (ⅲ) The PTT-induced ICD effect resulted in the formation of in situ whole-cell vaccines. (ⅳ) The enhanced antitumor immune efficiency was attributed to the immunomodulatory effect of MEL. Our findings elucidate the immunological aspects of combinatorial phototherapy and immune regulation, which will expand the scope of synergistic antitumor immunotherapy.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ying Sun: Writing – original draft, Methodology, Formal analysis, Data curation. Minglong Chen: Visualization, Formal analysis, Conceptualization. Ying Chen: Writing – original draft, Formal analysis, Data curation. Wanchen Zhao: Methodology, Formal analysis. Yanping Fu: Visualization, Data curation. Zhengwei Huang: Data curation, Conceptualization. Chao Lu: Visualization, Formal analysis. Chuanbin Wu: Software, Resources. Xin Pan: Resources, Methodology, Funding acquisition. Guilan Quan: Writing – review & editing, Supervision, Resources, Project administration, Funding acquisition, Conceptualization.
This study was supported by the National Natural Science Foundation of China (Nos. 82173747, 82373803), and the Special Fund Project for Science and Technology Innovation Strategy of Guangdong Province (No. 2021TQ060944).
Supplementary material associated with this article can be found, in the online version, at doi:
A.C. Huang, R. Zappasodi, Nat. Immunol. 23 (2022) 660–670. doi: 10.1038/s41590-022-01141-1
M. Saxena, S.H. van der Burg, C.J.M. Melief, N. Bhardwaj, Nature Rev. Cancer 21 (2021) 360–378. doi: 10.1038/s41568-021-00346-0
N. Coleman, M. Ameratunga, J. Lopez, Clin. Med. Insights Oncol. 12 (2018) 18.
Y. Iwai, J. Hamanishi, K. Chamoto, T. Honjo, J. Biomed. Sci. 24 (2017) 26.
R.C. Sterner, R.M. Sterner, Blood Cancer J. 11 (2021) 69.
J.S.W. Borgers, J. Haanen, Hematol. Oncol. Clin. North Am. 35 (2021) 129–144.
S.Y. Liu, W. Wei, H. Yue, et al., Biomaterials 34 (2013) 8291–8300.
Z. Hu, P.A. Ott, C.J. Wu, Nat. Rev. Immunol. 18 (2018) 168–182. doi: 10.1038/nri.2017.131
O.J. Finn, Nat. Rev. Immunol. 3 (2003) 630–641. doi: 10.1038/nri1150
B.S. Tian, R.X. Tian, S.H. Liu, et al., Adv. Mater. 35 (2023) 2304262.
D.V. Krysko, A.D. Garg, A. Kaczmarek, et al., Nat. Rev. Cancer 12 (2012) 860–875. doi: 10.1038/nrc3380
H. Monaco, S. Yokomizo, H.S. Choi, S. Kashiwagi, View-China 3 (2022) 22.
S.J. Park, W.D. Ye, R. Xiao, et al., Oral Oncol. 95 (2019) 127–135.
Z.L. Li, X.Q. Lai, S.Q. Fu, et al., Adv. Sci. 9 (2022) 2201734.
L. Yang, Z.Y. Zhao, B.S. Tian, et al., Nat. Commun. 15 (2024) 7499. doi: 10.1007/s11071-024-09375-4
K.F. Chu, D.E. Dupuy, Nat. Rev. Cancer 14 (2014) 199–208. doi: 10.1038/nrc3672
X.L. Yi, Q.Y. Duan, F.G. Wu, Research 2021 (2021) 9816594.
C.X. Zheng, X.X. Liu, Y.Y. Kong, et al., Acta Pharm. Sin. B 12 (2022) 3398–3409.
J.J. Zhu, R.M. Chang, B.L. Wei, et al., Research 24 (2022) 0295.
W.J. Yang, S.K. Yang, F. Zhang, et al., Ann. Transl. Med. 8 (2020) 399.
A. Mantovani, S. Sozzani, M. Locati, P. Allavena, A. Sica, Trends Immunol. 23 (2002) 549–555.
R. Noy, J.W. Pollard, Immunity 41 (2014) 49–61.
K. Matsuzaki, S. Yoneyama, K. Miyajima, Biophy. J. 73 (1997) 831–838.
K. Hristova, C.E. Dempsey, S.H. White, Biophy. J. 80 (2001) 801–811.
G. Gajski, V. Garaj-Vrhovac, Environ. Toxicol. Pharmacol. 36 (2013) 697–705.
C. Lyu, F.F. Fang, B. Li, Curr. Protein Pept. Sci. 20 (2019) 240–250. doi: 10.2174/1389203719666180612084615
C. Lee, H. Jeong, Y. Bae, et al., J. Immunother. Cancer 7 (2019) 147.
S. Kim, I. Choi, I.H. Han, H. Bae, Int. J. Mol. Sci. 23 (2022) 15751. doi: 10.3390/ijms232415751
X. Yu, Y.F. Dai, Y.F. Zhao, et al., Nat. Commun. 11 (2020) 1110.
Y. Chen, Q. Zeng, B. Chu, et al., Chin. Chem. Lett. 34 (2023) 108133.
D. Yang, M.L. Chen, Y. Sun, et al., Acta Biomater. 121 (2021) 119–133.
Y.Z. Chen, J.J. Zhu, J.S. Ding, W.H. Zhou, Chin. Chem. Lett. 35 (2024) 108706.
W. Li, J.Y. Chen, R.N. Terry, et al., J. Control. Release 347 (2022) 489–499.
Y.J. Huang, H.L. Lai, J.W. Jiang, et al., Asian J. Pharm. Sci. 17 (2022) 679–696. doi: 10.3390/agronomy12030679
N. Jiang, Z.G. Zhou, W. Xiong, et al., Chin. Chem. Lett. 32 (2021) 3948–3953.
D.J. Shen, L.H. Zhu, Y. Liu, et al., Nanotechnology 31 (2020) 18.
C.Y. Liao, M.E. Selvan, J. Zhao, et al., J. Phys. Chem. B 119 (2015) 10390–10398. doi: 10.1021/acs.jpcb.5b03254
W. Li, J. Yang, L.H. Luo, et al., Nat. Commun. 10 (2019) 245102.
H. Kobayashi, P.L. Choyke, Acc. Chem. Res. 52 (2019) 2332–2339. doi: 10.1021/acs.accounts.9b00273
C. Lee, S. Kim, C. Jeong, et al., Int. J. Mol. Sci. 23 (2022) 2157. doi: 10.3390/ijms23042157
M.Y. Qu, H.J. Kim, X.W. Zhou, et al., Nanoscale 12 (2020) 16724–16729. doi: 10.1039/d0nr02759f

Scheme 1 Schematic illustration of DMNs-assisted in situ cancer vaccine combined with cytolytic peptide for antitumor immunotherapy.

Figure 1 Preparation and characterization of MEL/ICG-HA. (A) Preparation process of MEL/ICG-HA. (B) TEM image and (C) particle size distribution of MEL/ICG-HA. Scale bar: 100 nm. (D) CD spectra of free MEL, MEL/ICG, and MEL/ICG-HA. (E) Ultraviolet and (F, G) fluorescence spectra of free ICG, MEL/ICG, and MEL/ICG-HA. (H) The temperature change curve of MEL/ICG-HA suspension under continuous NIR laser irradiation (808 nm, 1 W/cm2) for 5 min.

Figure 2 In vitro immune response of MEL/ICG-HA. (A) CLSM images of B16 cells staining by PI following treatment with free MEL and MEL/ICG-HA for 10 min. (B) CLSM images of B16 cells after incubation with free ICG and MEL/ICG-HA for different intervals. (C) Cell viability of B16 cells after treated with different concentrations of free MEL, ICG, and MEL/ICG-HA with or without laser irradiation. (D) CLSM images of CRT and HSP70 exposure in B16 cells after different treatments. (E, F) The flow cytometric analysis and quantification results of BMDCs and BMDMs. G1–G5 represent the cells incubated with B16 cells pretreated with (1) control, (2) MEL, (3) ICG + laser, (4) MEL/ICG-HA, and (5) MEL/ICG-HA + laser treatments. Scale bar: 20 µm. Data are presented as mean ± standard deviation (SD) (n = 3). **P < 0.01, ***P < 0.001, ****P < 0.0001.

Figure 3 Characterization of MEL/ICG-HA@DMNs. (A) Microscope image, (B) scanning electron microscope (SEM) image (scale bar: 400 µm), and (C) CLSM images of MEL/CY5.5-HA@DMNs (scale bar: 200 µm). (D) Layer-by-layer scanning of MEL/CY5.5-HA@DMNs (scale bar: 200 µm). (E) Images of gelatin skin model after application of MEL/ICG-HA@DMNs at various time points, the outline of needle tip is traced by the red lines (scale bar: 200 µm). (F) Image of gelatin following removal of MNs (scale bar: 200 µm). (G) Fluorescence signal distribution in gelatin skin model after MEL/CY5.5-HA@DMNs application (scale bar: 200 µm). MFI, mean fluorescence intensity.

Figure 4 (A) CLSM images of rat skin following application of MEL/ICG-HA@DMNs (scale bar: 100 µm). (B) Diffusion and penetration of MEL/ICG-HA@DMNs in the excised rat skin (scale bar: 200 µm). (C) Fluorescent images and (D) semiquantitative examination of B16-bearing mice following MEL/ICG-HA administration through DMNs and intratumor injection. (E) Thermal images and (F) maximum temperature change curve of tumor site under laser irradiation for 5 min after MEL/ICG-HA@DMNs administration. Data are presented as mean ± SD (n = 3). ****P < 0.0001.

Figure 5 In vivo antitumor efficacy and mechanism study: (A) Schematic diagram of the treatment protocol. Tumors were treated with DMNs and irradiated with a NIR laser (808 nm, 1 W/cm2) for 5 min at 1 h post-application. For intra-tumoral injection, MEL/ICG-HA were injected using insulin needles in a multipoint manner, and the administration site was irradiated with a NIR laser (808 nm, 1 W/cm2) for 5 min. (B) Tumor volume of mice after various treatments (n = 6). (C) H&E, Ki67, and TUNEL staining of tumor tissue after different treatments (scale bar: 100 µm). The quantification results of flow cytometric analysis of (D) CD80+CD86+DCs, (E) M1-type macrophage (F4/80+CD86+), (F) M2-type macrophage (F4/80+CD206+), (G) CD3+CD4+ T cells, (H) CD3+CD8+ T cells, and (I) CD45+CD3+ T cells in mice lymph node. G1: control; G2: MEL@DMNs; G3: MEL/ICG-HA@DMNs; G4: ICG@DMNs + laser; G5: MEL/ICG-HA@DMNs + laser; G6: MEL/ICG-HA@I.T. + laser (n = 3). ns, no significant difference. Data are presented as mean ± SD. P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载: