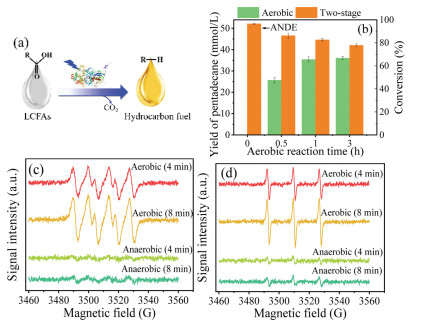
Figure 1.
Influence of oxygen on photoenzymatic decarboxylation. (a) Schematic diagram of CvFAP catalysis. (b) Inhibition of oxygen to CvFAP. (c) EPR spectra of DMPO/O2•−. (d) EPR spectra of TEMP/1O2.
Anaerobic environment as an efficient approach to improve the photostability of fatty acid photodecarboxylase
Xiaobo Guo , Ao Xia , Wuyuan Zhang , Feng Li , Yun Huang , Xianqing Zhu , Xun Zhu , Qiang Liao

The International Air Transport Association targeted to achieve net-zero carbon emission for the global air transport industry by 2050, implying 21.2 gigatonnes of carbon emission to be abated from 2021 to 2050 [1]. However, the aviation sector, particularly for long-haul flights, will not be electrification but will be highly dependent on liquid fuel for the foreseeable future [2]. It is necessary to incentivize the production of sustainable aviation fuel (SAF) from biomass waste such as waste cooking oil and lignocellulosic residues as alternative aviation fuel since the carbon reduction of SAF production is up to 90% compared to petroleum aviation fuel [2-7]. Unfortunately, harsh reaction conditions and low selection limit SAF production to less than 0.1% of the current global aviation fuel consumption [8, 9].
Fatty acid photodecarboxylase of Chlorella variabilis NC64A (CvFAP), a novel photoenzyme, has great potential in the decarboxylation of long-chain fatty acids (LCFAs) from waste cooking oil for SAF production since it is blue light-driven, irreversible, and cofactor-independent reaction (Fig. 1a) [10-12]. Furthermore, CvFAP, as a fascinating enzyme, has shown a promising application in energy chemistry, food chemistry, and organic synthesis [13-17]. However, the fragile nature that CvFAP is readily inactivated under blue-light illumination, especially in the absence of substrate, remains an urgent challenge [18-21]. Only two approaches were currently proposed to promote the stability of CvFAP, namely using crude enzyme containing CvFAP in place of the purified enzyme [18] and adding caprylic acid in the photoenzymatic decarboxylation [22]. However, CvFAP in the crude enzyme will also suffer from photoinhibition by high-intensity blue light, and the addition of caprylic acid will introduce impurity and compete for the active site of CvFAP with the target substrate.
The mechanism that blue light inhibits CvFAP, particularly in the absence of substrate, remains opaque [23]. It was reported that the singlet excited state of flavin adenine dinucleotide (1FAD*) decayed to the FAD ground state directly with fluorescence emission or to the metastable triplet state 3FAD* in the absence of substrate [10]; no inhibition of CvFAP was, however, reported under the blue-light illumination. A radical presumably derived from FAD semiquinone and protein-based organic radicals under blue-light illumination was previously reported to induce a modification of protein structure in or near the enzyme active site, thereby suppressing the activity of CvFAP after blue-light preillumination for 30 min [19]. Photoexcited flavin was suggested to oxidize active-site borne amino acid residues such as Cys432 or Arg451, thereby leading to an inactivation of CvFAP [22]. However, no specific radical accounting for the inhibition was detected in these works, and the inhibition mechanism of blue light to CvFAP remains to be explored.
Flavin readily undergoes photosensitization under blue-light illumination along with the generation of free radicals and radical ions via electron transfer and proton transfer, respectively, and the formation of singlet oxygen (1O2) via energy transfer [24-26]. Meanwhile, the generated free radicals can damage an amino acid residue of a flavoenzyme [24, 27, 28]. Inspired by that oxygen induces the generation of reactive oxygen species (ROS) in the photosensitization of flavoenzyme, we reasoned that oxygen played a vital role in the inhibition of flavoenzyme CvFAP under blue-light illumination. Thus, anaerobic environment to mitigate the production of ROS was hypothesized to facilitate a high photostability of CvFAP. Herein, the influence of oxygen on the photostability of CvFAP was investigated, and the inhibitory mechanism of CvFAP under blue-light illumination was explored.
To investigate the influence of oxygen on photoenzymatic decarboxylation, a two-stage decarboxylation of palmitic acid (54 mmol/L) was conducted, where the decarboxylation under aerobic environment (namely aerobic decarboxylation) for various times (i.e., 0, 0.5, 1, and 3 h) was first performed, which was subsequently transferred to anaerobic environment for another 3 h (namely anaerobic decarboxylation). For the sake of description, the anaerobic decarboxylation for 3 h without pre-aerobic decarboxylation was denoted as ANDE. As depicted in Fig. 1b, the yield of pentadecane in the aerobic decarboxylation for 3 h was 36.0 mmol/L. Interestingly, 52.1 mmol/L of pentadecane was obtained with the conversion of palmitic acid at 96.5% in the ANDE. Therefore, ANDE significantly improved the pentadecane yield by 44.7% compared to aerobic decarboxylation. In contrast, no difference in the pentadecane yield by CvFAP between aerobic environment and anaerobic environment (Ar, N2, or H2) was reported [18], which was due to that a pentadecane yield at 100% was obtained at a low substrate concentration (13 mmol/L) in 3 h and the difference was missed. Additionally, the conversion of palmitic acid increased from 47.5% to 66.7% after prolonging the aerobic decarboxylation time from 0.5 h to 3 h. The final conversion of palmitic acid after the two-stage decarboxylation decreased from 86.2% to 77.8% when increasing the aerobic decarboxylation time from 0.5 h to 3 h (Fig. 1b). The identical yield of pentadecane in aerobic decarboxylation for 1 and 3 h suggests that CvFAP was almost inactive in the aerobic decarboxylation for 1 h. However, pentadecane was once again produced after the assays were transferred to anaerobic environment, suggesting that oxygen induced a reversible inhibition to CvFAP but deactivated CvFAP after a long period of blue-light illumination. Analogously, ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) can be reversibly inhibited by oxygen [29].
The typical ROS such as superoxide radical (O2•−) and 1O2 at the initial 4 and 8 min of the photoenzymatic decarboxylation under aerobic or anaerobic environment were detected with electron paramagnetic resonance (EPR) to further explore the inhibition of oxygen to photoenzymatic decarboxylation [26, 30]. A specific TEMP/1O2 spin adduct giving rise to three resolved peaks and DMPO/O2•− spin adduct generating four resolved peaks were detected in aerobic decarboxylation (Figs. 1c and d). Intensities of both signals at 8 min were stronger than those at 4 min, suggesting increases of O2•− and 1O2 production in response to the reaction time from 4 min to 8 min. However, the weak signals of O2•− and 1O2 were detected in anaerobic decarboxylation, presumably due to little oxygen present after the degassing process (i.e., 75 ppm of O2 in the anaerobic chamber). O2•− and 1O2 generated from the photosensitization of riboflavin have been reported to damage amino acid residues in an intact flavoprotein, such as tyrosine, histidine, tryptophan, cysteine, and methionine [24, 27]. Therefore, the accumulation of O2•− and 1O2 in aerobic decarboxylation accounted for damage of ROS to amino acid residues in the active site of CvFAP, causing a decrease in the yield of pentadecane. Anaerobic environment could efficiently eliminate the accumulation of O2•− and 1O2 [31], thereby being beneficial to the CvFAP photostability.
Blue-light preilluminating CvFAP in the absence of substrate can severely inhibit the activity of CvFAP [18, 19, 22]. Herein, CvFAP BCs were firstly preilluminated under aerobic or anaerobic environment and then were supplemented with palmitic acid to assess the residual activity of CvFAP with anaerobic decarboxylation for 3 h. Preillumination under aerobic environment for 0.5 h caused a very low pentadecane yield (0.2 mmol/L) after the decarboxylation (Fig. 2a). Prolonging the aerobic preillumination time to 1 or 3 h, no pentadecane was produced after the decarboxylation, indicating an entire inactivation of CvFAP in the preillumination phase. Interestingly, after preilluminating CvFAP BCs under anaerobic environment for 0.5 h, 40.5 mmol/L of pentadecane was produced in the decarboxylation; the residual activity of CvFAP was 80.4%, which is 258.7 times higher than that of aerobic preillumination (Fig. 2b). When increasing the anaerobic preillumination time from 0.5 h to 3 h, the conversion of palmitic acid after the decarboxylation decreased from 74.9% to 47.4%. Consequently, an anaerobic environment significantly improved the CvFAP photostability in the preillumination with absent palmitic acid compared to aerobic environment.
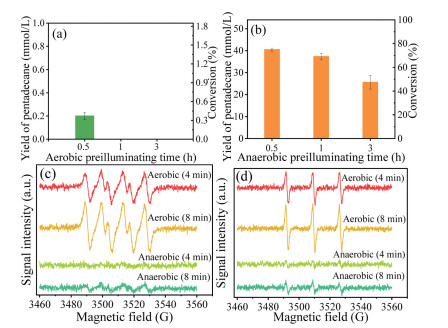
Remarkable specific signals of TEMP/1O2 spin adduct and DMPO/O2•− spin adduct were detected at the initial 4 and 8 min of the aerobic preillumination (Figs. 2c and d). Increases in O2•− and 1O2 production in response to the reaction time from 4 min to 8 min were also observed. The significant accumulation of O2•− and 1O2 in blue-light preillumination under an aerobic environment caused rapid inactivation of CvFAP in 30 min, thereby leading to almost no pentadecane produced after the decarboxylation (Fig. 2a). Weak signals of O2•− and 1O2 in anaerobic preillumination were also observed, which were derived from residual oxygen in the degassing process. Increasing the anaerobic preillumination time decreased the conversion of palmitic acid from 74.9% to 47.4% in the decarboxylation due to the suppression of CvFAP by little O2•− and 1O2 present in the preillumination (Fig. 2b). Moreover, the FAD semiquinone (FADH•) could play a vital role in the inhibition of CvFAP under anaerobic environment, as it was previously reported to be oxidative under an anaerobic environment [25, 32, 33]. Taken together, an anaerobic environment significantly improved the photostability of CvFAP under blue-light preillumination compared to that of aerobic preillumination. Various flavoproteins such as thioredoxin reductase and cryptochrome in E. coli were previously reported to be restrained by excessive blue-light exposure in the same pattern resemble to CvFAP, due to damages of O2•− and 1O2 to amino acid residues in a flavoprotein [27, 34-36]. Identically, inhibition of blue light to CvFAP was ascribed to the damage of O2•− and 1O2 generated in the photosensitization of FAD under aerobic environment to amino acid residues in or near the active site of CvFAP such as tyrosine and tryptophan [24]. Herein, significant signals of TEMP/1O2 spin adduct and DMPO/O2•− spin adduct were detected in the aerobic decarboxylation and anaerobic preillumination assay with E. coli broken cells containing an empty vector in place of the CvFAP BCs (Fig. S1 in Supporting information). Little ROS was detected in the anaerobic decarboxylation and aerobic preillumination assay with E. coli broken cells containing an empty vector (Fig. S1). Therefore, the source of O2•− and 1O2 could be derived from the photosensitization of other flavoproteins in addition to CvFAP in this study.
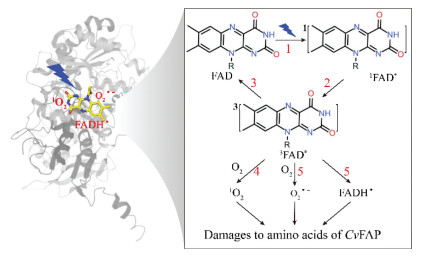
Given that CvFAP is a typical flavoprotein, the generation pathway of O2•− and 1O2 in photosensitization of CvFAP under aerobic environment can be proposed referring to that of riboflavin as depicted in Fig. 3 [31]. The FAD in CvFAP BCs is first excited by blue-light illumination to form a singlet excited state (1FAD*) (Step 1), which then undergoes rapid intersystem crossing to generate the triplet state 3FAD* (Step 2). Additionally, partial 1FAD* also enters the FAP photocycle for photoenzymatic decarboxylation. In terms of 3FAD*, one part can return to the ground state (Step 3), another part undergoes the Type-Ⅰ process (Step 4), and the third part enters the Type-Ⅱ process (Step 5). In the Type-Ⅰ process, O2•− and FADH• are produced along with an electron transfer from 3FAD* to O2 and a proton transfer to 3FAD*, respectively [33]. In the Type-Ⅱ process, 1O2 is produced after transferring the excitation energy of 3FAD* to O2 [37]. Consequently, the generated radicals such as O2•− and 1O2 under an aerobic environment and FADH• under anaerobic environment can damage amino acids in or near the active site of CvFAP, thereby causing a low photostability of CvFAP. Herein, l-cysteine was also used as a 3FAD* quencher in photoenzymatic decarboxylation, since l-cysteine was previously reported to efficiently quench the triplet excited state of riboflavin [38]. Addition of l-cysteine significantly improved the pentadecane yield in aerobic or anaerobic decarboxylation compared to the control (i.e., aerobic decarboxylation without l-cysteine addition). Pentadecane yield of anaerobic decarboxylation with l-cysteine addition increased by 34.6% compared to the control (Fig. S2 in Supporting information), indicating quenching 3FAD* beneficial to the high activity of CvFAP.
It was previously documented that CvFAP purified under ambient light conditions was readily inactive, but that purified under dim red light retained a high activity [19]. This is ascribed to that blue light in ambient light can induce the formation of O2•− and 1O2 in the photosensitization of FAD, but red light as well as a dark environment, cannot trigger the photosensitization. It was also previously demonstrated crude enzymes containing CvFAP enabled a higher yield of pentadecane even after a blue-light preillumination for 3 h than that of pure CvFAP [18]. Moreover, crude enzymes containing CvFAP exhibited a longer half-life time (ca. 19 h) than pure CvFAP (ca. 1 h) [22]. However, the crude enzymes will also be inhibited by the high intensity of blue light. The addition of caprylic acid impressively demonstrated an enhancement to the photostability of CvFAP, with the yield of pentadecane at 81% after preillumination for 24 h [22]. Caprylic acid helped to "keep the enzyme busy", thereby improving the photostability of CvFAP. It is noteworthy that this enhancement will fade away when caprylic acid is entirely consumed, and a high concentration of caprylic acid will compete for the active site of CvFAP with the target substrate [22]. In the present work, the photostability of CvFAP BCs resemble to the aforementioned crude enzyme was further improved under anaerobic environment. This is the third approach to improve the photostability of CvFAP to date.
In summary, anaerobic photodecarboxylation increased the conversion of palmitic acid by 44.7% compared to aerobic photodecarboxylation for 3 h. The residual activity of CvFAP was retained by 80.4% after anaerobic preillumination in the absence of palmitic acid for 0.5 h, which was 258.7 times higher than aerobic preillumination. Aerobic environment led to an accumulation of O2•− and 1O2 in the photosensitization of CvFAP, thereby suppressing the activity of CvFAP. Anaerobic environment eliminated the production of O2•− and 1O2, and efficiently improved the photostability of CvFAP. This study will facilitate the application of CvFAP in the utilization of waste lipids and the production of sustainable aviation fuel.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported by the National Natural Science Foundation of China (Nos. 52022015, 51876016), the Innovative Research Group Project of National Natural Science Foundation of China (No. 52021004), the State Key Program of National Natural Science of China (No. 51836001), the Chongqing Talents: Exceptional Young Talents Project (No. CQYC202005081), the Natural Science Foundation of Chongqing (No. cstc2021ycjh-bgzxm0160), and Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (No. TSBICIPCXRC-032).
Supplementary material associated with this article can be found, in the online version, at doi:
IATANet-Zero Carbon Emissions by 2050, International Air Transport Association, Canada, 2021 (accessed April 5, 2022). Available from
N.R. Baral, M. Yang, B.G. Harvey, et al., ACS Sustain. Chem. Eng. 9 (2021) 11872–11882. doi: 10.1021/acssuschemeng.1c03772
R. Shi, D.W. Archer, K. Pokharel, et al., ACS Sustain. Chem. Eng. 7 (2019) 18753–18764. doi: 10.1021/acssuschemeng.9b02150
Y.D. Chen, F.Y. Liu, N.Q. Ren, et al., Chin. Chem. Lett. 31 (2020) 2591–2602. doi: 10.1016/j.cclet.2020.08.019
K.W. Chew, J.Y. Yap, P.L. Show, et al., Bioresour. Technol. 229 (2017) 53–62. doi: 10.1016/j.biortech.2017.01.006
N. Kumar, C. Banerjee, J.S. Chang, et al., J. Clean. Prod. 362 (2022) 132114. doi: 10.1016/j.jclepro.2022.132114
Z. Deng, A. Xia, Q. Liao, et al., Biotechnol. Biofuels 12 (2019) 159. doi: 10.1186/s13068-019-1499-3
M. Li, J. Fu, P. Lv, et al., ACS Sustain. Chem. Eng. 8 (2020) 15484–15495. doi: 10.1021/acssuschemeng.0c03559
X. Yang, R.W. Jenkins, J.H. Leal, et al., ACS Sustain. Chem. Eng. 7 (2019) 14521–14530. doi: 10.1021/acssuschemeng.9b02041
D. Sorigue, B. Legeret, S. Cuine, et al., Science 357 (2017) 903–907. doi: 10.1126/science.aan6349
W. Zhang, M. Ma, M.M.E. Huijbers, et al., J. Am. Chem. Soc. 141 (2019) 3116–3120. doi: 10.1021/jacs.8b12282
D. Sorigue, K. Hadjidemetriou, S. Blangy, et al., Science 372 (2021) 148–159.
W. Zhang, J.H. Lee, S.H.H. Younes, et al., Nat. Commun. 11 (2020) 2258. doi: 10.1038/s41467-020-16099-7
M. Amer, E.Z. Wojcik, C. Sun, et al., Energy Environ. Sci. 13 (2020) 1818–1831. doi: 10.1039/D0EE00095G
H.J. Cha, S.Y. Hwang, D.S. Lee, et al., Angew. Chem. Int. Ed. 59 (2020) 7024–7028. doi: 10.1002/anie.201915108
F. Cheng, H. Li, D.Y. Wu, et al., Green Chem. 22 (2020) 6815–6818. doi: 10.1039/D0GC02696D
D. Li, T. Han, J. Xue, et al., Angew. Chem. Int. Ed. 60 (2021) 20695–20699. doi: 10.1002/anie.202107694
M.M.E. Huijbers, W. Zhang, F. Tonin, F. Hollmann, Angew. Chem. Int. Ed. 57 (2018) 13648–13651. doi: 10.1002/anie.201807119
B. Lakavath, T.M. Hedison, D.J. Heyes, et al., Anal. Biochem. 600 (2020) 113749. doi: 10.1016/j.ab.2020.113749
Y. Sun, E. Calderini, R. Kourist, ChemBioChem 22 (2021) 1833–1840. doi: 10.1002/cbic.202000851
X. Guo, A. Xia, F. Li, et al., Energy Convers. Manag. 255 (2022) 115311. doi: 10.1016/j.enconman.2022.115311
Y. Wu, C.E. Paul, F. Hollmann, ChemBioChem 22 (2021) 2420–2423. doi: 10.1002/cbic.202100182
D.J. Heyes, B. Lakavath, S.J.O. Hardman, et al., ACS Catal. 10 (2020) 6691–6696. doi: 10.1021/acscatal.0c01684
C.K. Remucal, K. McNeill, Environ. Sci. Technol. 45 (2011) 5230–5237. doi: 10.1021/es200411a
J. Pan, M. Byrdin, C. Aubert, et al., J. Phys. Chem. 108 (2004) 10160–10167. doi: 10.1021/jp037837b
F. Yang, X. Chu, J. Sun, et al., Chin. Chem. Lett. 31 (2020) 2784–2788. doi: 10.1016/j.cclet.2020.07.033
R.A. Lundeen, K. McNeill, Environ. Sci. Technol. 47 (2013) 14215–14223. doi: 10.1021/es404236c
C. Wu, Y. Li, Z. Cheng, et al., Chin. Chem. Lett. 33 (2022) 4339–4344. doi: 10.1016/j.cclet.2022.01.036
C. Bathellier, L.J. Yu, G.D. Farquhar, et al., Proc. Natl. Acad. Sci. U. S. A. 117 (2020) 24234–24242. doi: 10.1073/pnas.2008824117
X. Li, Y. Jia, J. Zhang, et al., Chin. Chem. Lett. 33 (2022) 2105–2110. doi: 10.1016/j.cclet.2021.08.054
D. Dolmans, F. Dai, R.K. Jain, Nat. Rev. Cancer 3 (2003) 380–387. doi: 10.1038/nrc1071
P. Zhang, P. Van Cappellen, K. Pi, et al., Environ. Sci. Technol. 54 (2020) 11622–11630. doi: 10.1021/acs.est.0c02916
M. Gauden, I.H.M. van Stokkum, J.M. Key, et al., Proc. Natl. Acad. Sci. U. S. A. 103 (2006) 10895–10900. doi: 10.1073/pnas.0600720103
R. Kern, A. Malki, A. Holmgren, et al., Biochem. J. 371 (2003) 965–972. doi: 10.1042/bj20030093
M. Insinska-Rak, M. Sikorski Chem, Eur. J. 20 (2014) 15280–15291. doi: 10.1002/chem.201403895
W. Li, X. Gong, X. Fan, et al., Chin. Chem. Lett. 30 (2019) 1775–1790. doi: 10.1016/j.cclet.2019.07.056
F. Xu, X.N. Song, G.P. Sheng, et al., ACS Sustain. Chem. Eng. 3 (2015) 1756–1763. doi: 10.1021/acssuschemeng.5b00267
D.R. Cardoso, D.W. Franco, K. Olsen, et al., J. Agric. Food Chem. 52 (2004) 6602–6606. doi: 10.1021/jf0401165

Figure 1 Influence of oxygen on photoenzymatic decarboxylation. (a) Schematic diagram of CvFAP catalysis. (b) Inhibition of oxygen to CvFAP. (c) EPR spectra of DMPO/O2•−. (d) EPR spectra of TEMP/1O2.

Figure 2 Influence of blue-light preillumination on decarboxylation reaction for 3 h. (a) Preillumination under aerobic environment. (b) Preillumination under anaerobic environment. (c) EPR spectra of DMPO/O2•− in preillumination. (d) EPR spectra of TEMP/1O2 in preillumination.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
