Citation:
Wang Wen, Liu Xiaolong, Chen Huan, Ling Ting, Xia Tian, Liu Xiumei, Liu Jing, Otkur Wuxiyar, Shi Xianzhe, Qi Huan, Chen Di, Piao Hai-Long. Cholesterol as a functional metabolite cooperates with metadherin in cancer cells[J]. Chinese Chemical Letters,
2020, 31(7): 1831-1834.
doi:
10.1016/j.cclet.2019.12.027
Cholesterol as a functional metabolite cooperates with metadherin in cancer cells
English
Cholesterol as a functional metabolite cooperates with metadherin in cancer cells
CAS Key Laboratory of Separation Science for Analytical Chemistry, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian 116023, China
b.
University of Chinese Academy of Sciences, Beijing 100049, China
* Corresponding author at: CAS Key Laboratory of Separation Science for Analytical Chemistry
Received Date:
24 October 2019 Accepted Date:
17 December 2019 Revised Date:
16 December 2019 Available Online:
15 July 2020
Abstract:
Protein-metabolite interactions (PMIs) play important roles in various biological processes, especially in disease progression. However, due to the complexity of living cells, it is very difficult to identify specific PMIs. Herein, we chose one oncogenic factor, metadherin (MTDH), as a bait to identify its in vivo interacting metabolites in cancer cells. Cholesterol is an important metabolite and essential structural component of cell membranes. It could also drive several diseases including cancer. Interestingly, we found that cholesterol robustly interacted with MTDH and downregulated the expression of MTDH in cancer cells. Furthermore, MTDH disturbed metabolite alterations under cholesterol treatment in MTDH transduced cancer cells. Collectively, our results uncover an undescribed PMI where MTDH, as an oncogenic factor, might positively regulate cancer progression by interacting with cholesterol. This study interprets the theoretical basis of PMI-oriented cancer progression and targeting therapies in clinic.
Interactions between proteins and metabolites are very important during the fundamental cellular signal transduction.Metabolites such as succinate [1], lactate [2] and fumarate [3] can either act as global regulators of many proteins or directly target on specific proteins in disease progression. In order to identify these key PMIs, several invitro and in vivo methods have been developed based on chemical communications [4]. Previously, methods based on immobilizing recombinant proteins and affinity chromatography were used to profile the nuclear receptor of PPARγ [5] and ERRα interacting lipidome ligands, in which cholesterol was identified to interact with ERRα significantly [6]. This affinity chromatography approach is widely applicable to detect ligands of receptor proteins, but these methods still give rise to non-specific metabolite-protein bindings which may disrupt the true PMIs.
Activity-based protein profiling (ABPP) is a chemical proteomic method based on clickable and photoreactive active probes [7-9]. Recently, ABPP is emerging as one of the most popular methods to identify new drug targets [10,11]. Meanwhile, several studies had identified many small metabolites interacting proteins through ABPP, such as palmitoylated proteins [12], lipid-binding proteins [13], ADP-ribosylation of GTPases [14], and cholesterylation of Smoothened [15]. Another chemoproteomic strategy combines limited proteolysis (LiP) with mass spectrometry in the presence of unmodified metabolites to systematically identify PMIs in Escherichia coli (E. coli) [16]. Although these methods have defined numerous interactions, these PMIs cannot represent the realistic cellular PMI landscape yet.
Herein, we are particularly interested in the oncogenic factor metadherin (MTDH), which was reported to play crucial roles in cancer progression [17]. To identify MTDH interacting metabolites, we modified the sensitive and scalable affinity purification method [18] to directly identify PMIs in mammalian cells and found the cholesterol enrichment and binding toMTDH.Thismodified strategy led to a functional binding between cholesterol and MTDH.
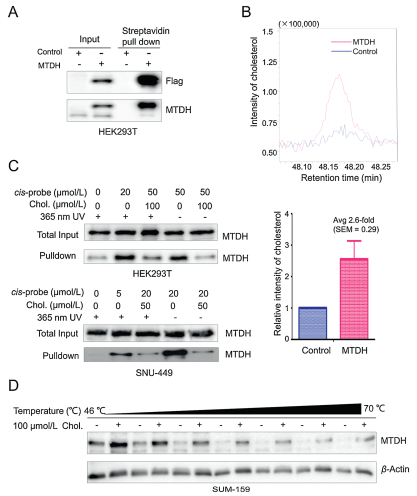
To identify MTDH interacting metabolites, we performed the activity-based metabolite profiling (ABMP) which is based on affinity protein purification and mass spectrometry analysis (Fig. S1 in Supporting information). We first transduced S-protein fused MTDH and control vector into HEK293T cells [19]. Compared with control cells, the levels of MTDH were significantly enriched in the MTDH transduced cells by S-beads pulldown purification (Fig. 1A). Subsequently, through the GC–MS-based untargeted metabolomics profiling, we found that there existed differential metabolites between these two pull-downed samples (Fig. S2A in Supporting information). Interestingly, we observed a 2.6-fold increase in the content of cholesterol by MTDH purification (Fig. 1B and Fig. S2A).
Figure 1
Figure 1.
Identification and validation of MTDH-cholesterol interaction. (A) SFBControl or SFB-MTDH was transfected into HEK293T cells. MTDH was purified with Streptavidin beads (S-beads) and immunoblotted with antibodies against Flag and MTDH. (B) Relative enrichment intensity of cholesterol from the control or SFBMTDH transfected cells. (C) HEK293T and SNU-449 cells were incubated with a cissterol probe or cholesterol and then irradiated with or without 365-nm UV light. After the pre-clear and CuAAC click reaction, samples were immunoblotted with antibody against MTDH. (D) Increased thermal stability of MTDH caused by the binding with cholesterol in SUM-159 cells was measured by the temperaturedependent cellular thermal shift assays. Data in (B) is presented as mean ± SEM. The experiments were repeated five times. Chol., cholesterol. See also Fig. S2 in Supporting information.
Next, we validated the specificity of the interaction between MTDH and cholesterol by the ABPP method [20]. HEK293T and SNU449 cells were treated with a cis-sterol probe in the absence or presence of excess cholesterol, respectively (Fig. S2B in Supporting information). Then we irradiated the cells with or without ultraviolet (UV) light for cross-linking to catch probe-interacting proteins, conjugated probe-interacting proteins to an azide-biotin tag by azide-alkyne cycloaddition (CuAAC) and enriched these proteins using streptavidin. We found that the cis-sterol probe specifically interacted with MTDH in HEK293T and SNU449 cells with or without UV irradiation, whereas these interactions were inhibited by exogenous cholesterol treatment (Fig. 1C). Furthermore, we performed cellular thermal shift assay to evaluate the thermal stabilization of MTDH upon cholesterol binding [21]. The results showed that cholesterol treatment increased the thermal stability of MTDH in SUM159, MHCC97H and HCC1973 cells (Fig. 1D and Fig. S2C in Supporting information). Taken together, these results suggested that cholesterol may directly interact with MTDH in different cells.
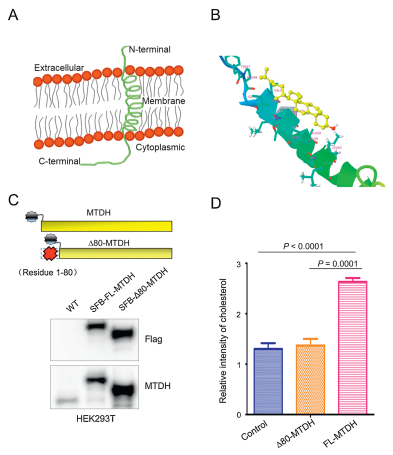
To dissect the cholesterol interaction region of MTDH, we next performed a computational docking analysis between cholesterol and MTDH based on their structural features. The MTDH sequence (Q86UE4) was obtained from UniProt Knowledgebase (https://www.uniprot.org/). Then, we used an online tool Phyre2 (www.sbg.bio.ic.ac.uk/phyre) to predict and analyze the protein structure of MTDH [22]. As a result, a transmembrane alpha-helix was predicted at the N-terminal residues (49–74) of MTDH (Fig. 2A). Considering the possible cholesterol-MTDH interaction in the membrane bilayer and frequent alpha-helix structures at ligandbinding domains driving protein activation [23], we carried out docking calculations between cholesterol and the N-terminal residues (49–74) of MTDH using DockingServer [24] to obtain cholesterol and MTDH binding information. Interestingly, hydrophobic interactions between cholesterol and the alpha-helix segment of MTDH lead to a 90% of prediction binding frequency (Fig. 2B). Furthermore, to investigate the importance of the MTDH alpha-helix segment for cholesterol interaction, we deleted the 1–80 N-terminal residues of MTDH (SFB-Δ80-MTDH) (Fig. 2C) and repeated GC–MS-based cholesterol detection analysis of purified MTDH-metabolite complexes in HEK293T cells transduced with control vector, SFB-MTDH and SFB-Δ80-MTDH (Figs. S3A and S3B in Supporting information). We observed that there were no significant differences in cholesterol contents between samples from the control vector and the SFB-Δ80-MTDH transduced cells (Fig. 2D), whereas significant enrichments of cholesterol were observed in the SFB-FL-MTDH transduced samples. Recently, Winkler et al., demonstrates that Niemann-Pick type C (NPC) proteins interact with cholesterol through its N-terminal alpha-helix membrane region in eukaryotic cells [25]. These data indicated that cholesterol may bind to MTDH through the transmembrane alpha-helix at N-terminal region.
Figure 2
Figure 2.
The N-terminal alpha-helix transmembrane domain of MTDH interacts with cholesterol. (A) The transmembrane alpha-helix domain was predicted at residues 49–74 of MTDH using Phyre2. (B) The binding mode between cholesterol (yellow) and MTDH (rainbow). This binding mode is calculated by DockingServer based on the structure information of cholesterol and the alpha-helical segment of MTDH. (C) Upper panel: schematic representation of the full length MTDH and N-terminal alpha-helix excluded MTDH. Lower panel: HEK293T cells were transfected with the SFB-tagged full-length MTDH or fragment of D80-MTDH. MTDH and the fragment were immunoblotted with antibodies against Flag. (D) Relative intensity of cholesterol enrichment from the control, SFB-Δ80-MTDH and SFB-FL-MTDH transfected cells. See also Fig. S3 in Supporting information. Data in (D) is presented as mean ± SEM. Statistical significance was determined by a two-tailed, unpaired Student's t-test. The experiments were repeated three times. FL, full length.
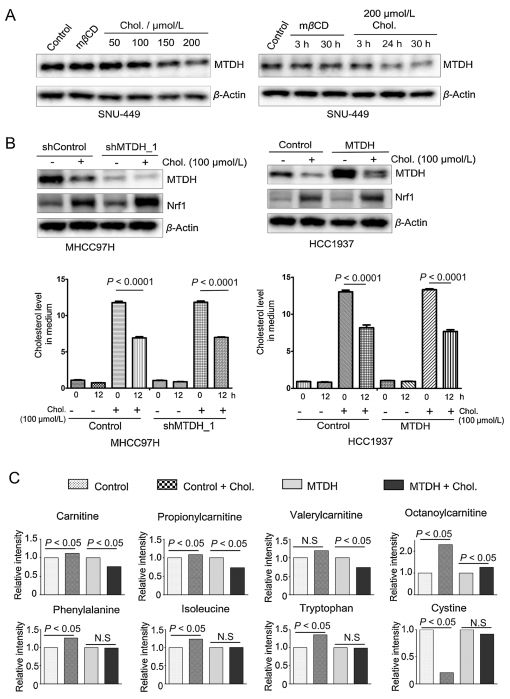
Cholesterol overload induces disease progression, especially for hepatocellular carcinoma [26]. To investigate whether cholesterol overload affects MTDH expression, we treated cells with or without cholesterol in a dose and time-dependent manner. Cholesterol significantly decreased MTDH expression in a dose and time-dependent manner compared with vehicle treatment (Fig. 3A and Fig. S4A in Supporting information). However, the nuclear factor erythroid 2 related factor-1 (Nrf1), a transcription factor that regulates cellular growth through metabolic alterations [27] and a positive response to cholesterol treatment [28], was significantly increased in both the MTDH knock-downed and overexpressed cells (Fig. 3B upper panel). In addition, the consumption of cholesterol was significantly increased in either the control or MTDH engineered cells compared with vehicle treatment cells (Fig. 3B lower panel). Furthermore, we found that cholesterol treatment could not turnover the nuclear localized MTDH but decreased the expression of MTDH (Fig. S4B in Supporting information). Taken together, the cholesterol-MTDH interaction can influence the MTDH stability.
Figure 3
Figure 3.
Identification of cholesterol as a functionally interacting metabolite of MTDH. (A) Immunoblotting of MTDH and β-actin in SNU449 cells with cholesterol or mβCD treatment. The cells were treated time and dose-dependently, respectively. (B) Upper panel: Immunoblotting of MTDH, Nrf1 and β-actin in the MTDH shRNA-transduced MHCC97H cells and the MTDH overexpressed HCC1937 cells, with or without cholesterol treatment. Lower panel: Quantification of cholesterol levels in cell culture medium after 12 h culture (n = 6). Statistical significance was determined by a two-tailed, unpaired Student's t-test. (C) The levels of carnitine, propionyl-carnitine, valeryl-carnitine, octanoyl-carnitine, phenylalanine, isoleucine, tryptophan and cystine in the control vector or MTDH transduced cells under cholesterol treatment (n = 6). Statistically significance of metabolites: P < 0.05 by two-way ANOVA. Data in (B) and (C) are presented as mean ± SEM. h, hours; Chol., cholesterol; N.S, not significant. See also Fig. S4 in Supporting information.
Metabolic adaptations are essential for cellular homeostasis in cancer. To dissect the causative link between cellular cholesterol homeostasis and MTDH, we performed GC–MS-based metabolic profiling in MTDH overexpressed or knock-downed cells. However, the cholesterol level was not significantly altered in the MTDH overexpressed or knock-downed cells (Fig. S4C in Supporting information). Next, to identify cellular metabolisms regulated by cholesterol-MTDH interactions, we performed liquid chromatography-mass spectrometry (LC–MS) based metabolic profiling in control or MTDH overexpressed HCC1937 cells with or without cholesterol treatment. We observed that amino acids, free carnitine, lipids, and several acyl-carnitines were significantly altered under the cholesterol treatment (Fig. S4D in Supporting information). Meanwhile, carnitine, propionyl-carnitine, valeryl-carnitine, octanoyl-carnitine, phenylalanine, isoleucine, methionine, tyrosine, lysine and tryptophan were increased in the control cells by the cholesterol treatment, whereas these metabolites in the MTDH overexpressed cells were not significantly influenced or even decreased with the cholesterol treatment. In contrast, the cystine and calcifediol were remarkably decreased in the control cells compared with the MTDH overexpressed cells (Fig. 3C and Fig. S4D). Collectively, MTDH-cholesterol interaction may affect MTDH stability, and regulates cellular metabolic reprograming in cancer cells.
Given the fact that most proteins are surrounded by various metabolites, investigation and profiling of in vivo active proteinmetabolite interactions are critical for understanding the molecular regulatory mechanisms in cells. Even though several studies had presented the protein-metabolite interaction in different cellular systems including E. coli [16] and mammalian cells [20], there still exists challenges for the systematic and high-throughput PMIs identification at physiological conditions. The non-covalent or hydrophilic metabolite-MTDH interactions were likely lost during the affinity purification and aqueous washes, due to this reason the new active chemical metabolite probes should be developed. Moreover, instead of the GC–MS-based profiling of limited thermally stable compounds, a universal small metabolite detection method is needed for untargeted analysis herein. With the development of mass spectrometry and modifications on purification, endogenous PMIs will be globally identified, which provides more insights into biological mechanistic networks.
In conclusion, this study demonstrated and identified active protein-metabolite binding complexes through the affinity purification and followed by the extraction of metabolites. Additionally, we integrated other chemobiological strategies to validate the MTDH-cholesterol binding and defined the functional role of MTDH-cholesterol interaction in cancer cells through the LC–MSbased metabolomics. The MTDH-cholesterol interaction inhibited the expression of MTDH; however, the detailed biological molecular mechanism should be defined in the future. But, this cholesterolMTDH interaction sheds light on the theoretical basis of PMI-oriented targeting treatment in clinic.
Declaration of competing interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank all members of the Dr. Piao laboratory for helpful discussions and suggestions. This study is supported by the National Natural Science Foundation of China (Nos. 81672440, 21575142), Innovation Program of Science and Research from the DICP, CAS (No. DICP ZZBS201803).
M.B.L. Winkler, R.T. Kidmose, M. Szomek, et al., Cell 179(2019) 485-497. doi: 10.1016/j.cell.2019.08.038
[26]
C. Enriquezcortina, O. Bellomonroy, P. Rosalescruz, et al., Oncotarget 8(2017) 104136-104148. doi: 10.18632/oncotarget.22024
[27]
T. Tsujita, V. Peirce, L. Baird, et al., Mol. Cell. Biol. 34(2014) 3800-3816. doi: 10.1128/MCB.00110-14
[28]
S.B. Widenmaier, N. Snyder, T. Nguyen, et al., Cell 171(2017) 1094-1109. doi: 10.1016/j.cell.2017.10.003
Figure 1
Identification and validation of MTDH-cholesterol interaction. (A) SFBControl or SFB-MTDH was transfected into HEK293T cells. MTDH was purified with Streptavidin beads (S-beads) and immunoblotted with antibodies against Flag and MTDH. (B) Relative enrichment intensity of cholesterol from the control or SFBMTDH transfected cells. (C) HEK293T and SNU-449 cells were incubated with a cissterol probe or cholesterol and then irradiated with or without 365-nm UV light. After the pre-clear and CuAAC click reaction, samples were immunoblotted with antibody against MTDH. (D) Increased thermal stability of MTDH caused by the binding with cholesterol in SUM-159 cells was measured by the temperaturedependent cellular thermal shift assays. Data in (B) is presented as mean ± SEM. The experiments were repeated five times. Chol., cholesterol. See also Fig. S2 in Supporting information.
Figure 2
The N-terminal alpha-helix transmembrane domain of MTDH interacts with cholesterol. (A) The transmembrane alpha-helix domain was predicted at residues 49–74 of MTDH using Phyre2. (B) The binding mode between cholesterol (yellow) and MTDH (rainbow). This binding mode is calculated by DockingServer based on the structure information of cholesterol and the alpha-helical segment of MTDH. (C) Upper panel: schematic representation of the full length MTDH and N-terminal alpha-helix excluded MTDH. Lower panel: HEK293T cells were transfected with the SFB-tagged full-length MTDH or fragment of D80-MTDH. MTDH and the fragment were immunoblotted with antibodies against Flag. (D) Relative intensity of cholesterol enrichment from the control, SFB-Δ80-MTDH and SFB-FL-MTDH transfected cells. See also Fig. S3 in Supporting information. Data in (D) is presented as mean ± SEM. Statistical significance was determined by a two-tailed, unpaired Student's t-test. The experiments were repeated three times. FL, full length.
Figure 3
Identification of cholesterol as a functionally interacting metabolite of MTDH. (A) Immunoblotting of MTDH and β-actin in SNU449 cells with cholesterol or mβCD treatment. The cells were treated time and dose-dependently, respectively. (B) Upper panel: Immunoblotting of MTDH, Nrf1 and β-actin in the MTDH shRNA-transduced MHCC97H cells and the MTDH overexpressed HCC1937 cells, with or without cholesterol treatment. Lower panel: Quantification of cholesterol levels in cell culture medium after 12 h culture (n = 6). Statistical significance was determined by a two-tailed, unpaired Student's t-test. (C) The levels of carnitine, propionyl-carnitine, valeryl-carnitine, octanoyl-carnitine, phenylalanine, isoleucine, tryptophan and cystine in the control vector or MTDH transduced cells under cholesterol treatment (n = 6). Statistically significance of metabolites: P < 0.05 by two-way ANOVA. Data in (B) and (C) are presented as mean ± SEM. h, hours; Chol., cholesterol; N.S, not significant. See also Fig. S4 in Supporting information.

 DownLoad:
DownLoad:

 下载:
下载:




