Citation:
Xiangning Fang, Qinglong Qiao, Fei Deng, Zhaochao Xu. Designing malachite green derivatives to optimize fluorogen-activating protein pairs for rapid PAINT super-resolution imaging in living cells[J]. Chinese Chemical Letters,
2025, 36(12): 110980.
doi:
10.1016/j.cclet.2025.110980
Designing malachite green derivatives to optimize fluorogen-activating protein pairs for rapid PAINT super-resolution imaging in living cells
English
Designing malachite green derivatives to optimize fluorogen-activating protein pairs for rapid PAINT super-resolution imaging in living cells
Received Date:
11 January 2025 Accepted Date:
19 February 2025 Revised Date:
18 February 2025 Available Online:
15 December 2025
Abstract:
Fluorogen-activating proteins (FAPs) selectively bind to specific fluorophores, inducing fluorescence activation through the inhibition of torsion of fluorophores. This binding-activation mechanism provides a highly specific and efficient fluorescence system that minimizes background signals, significantly enhancing the signal-to-noise ratio (SNR) and making it a powerful tool in live-cell imaging. The principle of binding-activation fluorescence is fundamental to point accumulation for imaging in nanoscale topography (PAINT) super-resolution imaging. However, the high binding affinity between traditional FAP-fluorophore pairs limits their application in PAINT, thus hindering the rapid and dynamic imaging necessary for high-resolution cellular studies. In this work, we designed malachite green (MG) derivatives with bulky N-substituents to modulate the binding affinity of the MG-dL5** fluorophore-FAP pair. This modification introduces steric hindrance in MG-dL5** system, resulting in reduced binding affinity and practicability for fast, high-resolution PAINT imaging. Among the synthesized derivatives, MG-Pen showed optimal properties, enabling rapid and high-resolution PAINT imaging of dL5** in living cells. This study highlights the potential of MG derivatives optimization in overcoming the limitations of fluorophore-FAP pairs for super-resolution imaging and provides a new approach for enhancing the performance of PAINT in living cell applications.
The rapid development of super-resolution fluorescence imaging techniques has revolutionized our ability to explore cellular structures and dynamics at the nanoscale [1–8]. Among these techniques, single-molecule localization microscopy (SMLM) has emerged as a powerful tool that overcomes the diffraction limit of light, enabling imaging of subcellular features with unprecedented resolution [9,10]. SMLM techniques, such as photoactivated localization microscopy (PALM) and stochastic optical reconstruction microscopy (STORM), rely on the stochastic activation and blinking of fluorescent probes [11–13], have achieved precise localization of individual fluorophores. Point accumulation for imaging in nanoscale topography (PAINT) is a subtype of SMLM that utilizes transient binding interactions between fluorophores and targets to achieve super-resolution imaging [14,15]. PAINT imaging achieves high resolution by accumulating localized points of fluorescence emission from sparsely and reversibly labelled targets [16,17].
PAINT requires sparse and reversible labeling to satisfy stochastic switching between fluorescent on and off state, which is essential for breaking the diffraction limit of light (Fig. 1a). This sparse binding minimizes the impact of the fluorescent probes on the target, thereby preserving the target's normal physiological functions. The reversibility of binding is crucial for maintaining the dynamic nature of the imaging process and preventing photobleaching, enabling continuous and high-resolution imaging of molecular interactions [18,19]. To satisfy the optimal binding-dissociation kinetics necessary for PAINT imaging, fluorophores with low affinity for the target are preferred [16]. Such fluorophores facilitate rapid and sparse fluorescent on-off switch, which is the key to stochastic super-resolution imaging.
Figure 1
Figure 1.
Mechanism of PAINT imaging and MG-dL5** imaging system. (a) Process and requirement of PAINT imaging. (b) Binding of MG and dL5** lead to fluorescence activation. (c) Molecule design of MGs with bulky N-substituents to achieve rapid PAINT imaging.
Since Hochstrasser group first demonstrated PAINT imaging of lipid membranes using Nile Red in 2006 [14], various PAINT-based techniques have been developed for different applications. For example, DNA-PAINT utilize programmable complementary base pairing for target labeling [20,21], and Glyco-PAINT leverage the interaction between glycans and lectins for lectin imaging [22]. These techniques have expanded the scope of PAINT to include quantitative analysis, multiplexing, and single-molecule analysis [23–25]. More recently, fluorogen-activating proteins (FAPs) that trigger fluorescence emission upon binding to specific fluorophores have gained attention [26–29]. Fluorophore-FAP pairs with reversible binding properties have proven effective for PAINT imaging, offering new avenues for high-resolution live-cell imaging.
Among the various fluorophore-FAP pairs, the malachite green-dL5** system has become one of the most widely used pairs due to the high brightness and near-infrared emission of malachite green (MG) (Fig. 1b). Since Szent-Gyorgyi group developed the MG-dL5** system in 2008 [30], many groups have applied it to a variety of targets labeling, and expanded a wide range of biological imaging applications [31–33]. The FAP dL5** could be easily fused to target protein as a tag, such as nuclear localization sequence (NLS)-myc in nuclei, cyclooxygenase Ⅳ-cyclooxygenase 8 (COXⅣ-COX8) in mitochondria and Lys-Asp-Glu-Leu (KDEL) in endoplasmic reticulum. And the MG probes could specifically bind with dL5** to activate fluorescence for labeling and imaging. Before binding with dL5**, MG probes are fluorescence quenching due to the twisted intramolecular charge transfer (TICT) effect caused by the torsion of C—N bond of the conjugated system. MG probes emit strong fluorescence after embedding the binding cavity of dL5**, thus could achieve no-wash imaging. The binding between MG and dL5** is highly specific, and can be completed in several minutes, hence the MG-dL5** system has become a widely used tool in fluorogenic imaging. Although the non-covalent binding between MG and dL5** is reversible, the high binding affinity between the two components is inappropriate for efficient PAINT imaging. Bruchez group addressed this challenge by precisely controlling the labeling density [34]. They used extremely low MG concentrations (picomolar level) and dye titration to achieve sparse labeling of dL5**. They also adjusted the acquisition cycle and acquisition time to wait for the bleached dyes to be replaced by fresh dyes to restore the labeling. Through the precise operation during imaging, they finally achieved high-precision PAINT imaging of actin in living cells. However, this complex method requires nearly 1 h to complete one PAINT image, which limits real-time and dynamic living cell imaging. Consequently, it is an important challenge to accelerate the speed of PAINT imaging utilizing MG-dL5** system.
In this study, we developed a series of MG derivatives designed to optimize the MG-dL5** system for rapid PAINT imaging (Fig. 1c). Rather than the traditional method of modifying FAP, we introduced bulky N-substituents on MG to alter the binding affinity for dL5**. As the traditional N-methyl MG (MG-TM) binding to dL5**, the four carbons of the methyl group occupy half of the contact area within the dL5** binding cavity [35]. Hence we hypothesized that introducing N-substituents with larger volume in MG would increase steric hindrance in MG-dL5** system, thus reducing binding affinity and satisfying the required binding-dissociation kinetics for PAINT imaging. The volume of azacyclic alkane is larger than the traditional N-methyl, and the rigid ring is conducive to inhibiting TICT and improving the brightness of MGs. Therefore, we used azetidine, azetidine-3-ol, azetidine-3-fluoro and azacyclopentane as N-substituents of MG, and synthesized MG-Aze, MG-Aze-OH, MG-Aze-F and MG-Pen, respectively. Among these derivatives, MG-Pen showed optimal characteristics, enabling rapid PAINT imaging in living cells in just 2 min, without the need for careful labeling density control. This innovation expands the potential of FAP-based systems for super-resolution imaging, providing a powerful tool for real-time, high-resolution imaging of dynamic molecular processes in living cells.
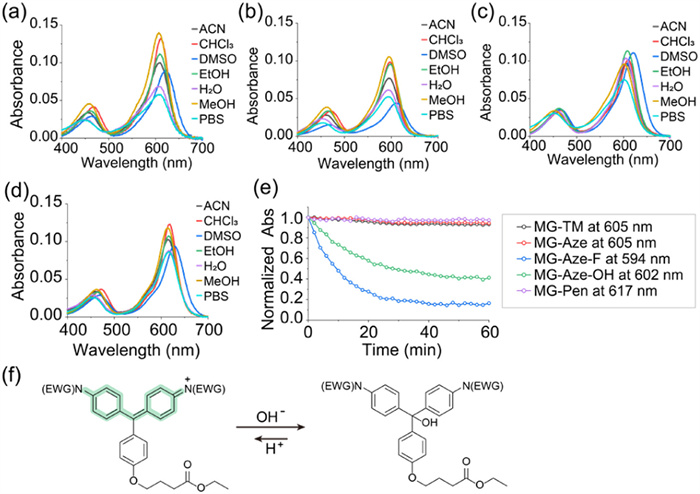
The synthesis of MG-Aze, MG-Aze-OH, MG-Aze-F and MG-Pen were successfully accomplished from bisphenol compounds using Pd-catalyzed C—N cross-coupling reactions (Scheme S1 in Supporting information). We first investigated the photophysical properties of these MG derivatives with bulky N-substituents (Figs. 2a–d, Fig. S1 and Tables S1–S4 in Supporting information). Each MG derivative showed two absorbance bands, the main absorbance band with long wavelength at approximately 600 nm was formed by the two N-substituents and the connected benzene rings, and the secondary absorbance band with blue-shifted wavelength at around 450 nm was formed by one N-phenyl and the orthogonal, pendant ring [36]. The maximum absorption wavelength of MG-Aze is 605 nm, which is consistent with the traditional MG-TM. The maximum absorption of MG-Aze-OH is blue-shifted to 602 nm due to the electron-withdrawing effect of OH group, and the fluorine with a stronger electron-withdrawing effect causes further blue-shifted of MG-Aze-F to 594 nm, while the maximum absorption wavelength of MG-Pen is red-shifted to 617 nm due to the electron-donating effect of cyclopentane. In addition, these four MGs were fluorescence quenching in solutions due to the strong TICT effect caused by the torsion of the conjugated system.
Figure 2
Figure 2.
Photophysical properties of MGs with bulky N-substituents. (a–d) Ultraviolet–visible (UV–vis) spectra of MG-Aze (a), MG-Aze-F (b), MG-Aze-OH (c) and MG-Pen (d) in different solvents. (e) The fluctuation of normalized absorbance of MG-TM, MG-Aze, MG-Aze-F, MG-Aze-OH and MG-Pen in water during 1 h. (f) Mechanism of MG-Aze-F and MG-Aze-OH bleaching in water. The concentration of sample is 2 µmol/L.
During the experiment of photophysical properties, an interesting phenomenon was observed. In aqueous solutions, MG-Aze-OH and MG-Aze-F showed rapid bleaching, that the solution gradually changed from green to nearly colorless within a few minutes. To further explore the abnormal phenomenon, we monitored the absorption spectrum of these MGs in water every 2 min for 1 h (Fig. 2e and Fig. S2 in Supporting information). During the spectrum monitoring, the absorbance of MG-TM, MG-Aze, and MG-Pen remained nearly constant, while the absorbance of MG-Aze-OH and MG-Aze-F decreased rapidly within the initial 20 min and reached equilibrium at 40 min. The absorbance of MG-Aze-OH at 602 nm decreased to 40% of the initial absorbance, and the absorbance of MG-Aze-F at 594 nm decreased more drastically to 15% of the initial value.
The hydroxyl group and fluorine group have strong electron-withdrawing effect, hence the modification of azetidine-3-ol and azetidine-3-fluoro to replace N-methyl would lead to the electron deficiency of MG's meso‑carbon. Therefore, the electron-donating hydroxyl in water would attack the meso‑carbon of MG-Aze-OH and MG-Aze-F and undergo nucleophilic additive reaction, resulting in the products with a destroyed conjugated system, which reduce the absorbance of the MGs in water (Fig. S3 in Supporting information). Meanwhile, protons in water make the nucleophilic reaction reversible, and eventually reach to the dynamic equilibrium, make the absorbance eventually remain stable (Fig. 2f). The electron-withdrawing effect of fluorine group is more severe than that of hydroxyl group, so the absorbance of MG-Aze-F decreased faster and more acutely than that of MG-Aze-OH, making it easier to be bleached in water. MG-Aze-OH and MG-Aze-F are not suitable for practical applications of labeling and imaging in living cells due to the instability in water. However, MG-Aze and MG-Pen could exist stably in water and have the potential for dL5** labeling and imaging in living systems.
We subsequently performed molecular docking of MG-Aze and MG-Pen with dL5**, to evaluate the binding affinity of dyes and protein. We constructed the three-dimensional (3D) structure of dL5** (D50E mutation) by homology modeling based on the MG-TM-bound L5* (PDB ID: 4K3H). As shown in Figs. 3a–c, MG-Aze and MG-Pen were located in the binding cavity formed by the symmetrical antiparallel homotypic L5* protein dimer (dL5**), inserted in the slot formed by the side-chain of Tyr96, with the pendent ring extending outward from the cavity, which were almost consistent with the performance of MG-TM.
Figure 3
Figure 3.
Binding affinity evaluation of MGs. (a–c) Molecular docking analysis of MG-Aze (a), MG-Pen (b) and MG-TM (c). Left: frontal view of MGs/dL5** complex and calculated affinity; middle: spatial distribution of amino acid side chains in dL5** that contact MGs; right: electric charge density of MGs and binding cavity. (d) The twisting analysis of azacyclopentane of MG-Pen binding with dL5**. (e) Confocal imaging of Hela cells co-expressing mCer3 and dL5** in nuclei incubated with MGs (500 nmol/L). Scale bar: 10 µm.
The calculated binding affinity of docking MG-dL5** complex for MG-Aze is −8.6 kcal/mol, which is similar to the traditional MG-TM (−9.5 kcal/mol). However, MG-Pen with larger N-substituent performed lower calculated binding affinity as −3.5 kcal/mol, which has decreased by about two-thirds compared with MG-TM. The charge density pattern showed that the electron cloud of MG-TM and MG-Aze had nearly no contact with the electron cloud of amino acid residues in the binding site, while the electron cloud of the azacyclopentane of the MG-Pen exceeded the binding cavity and overlapped with the electron cloud of the amino acid residues.
Bruchez group has analyzed the interaction between MG-TM and amino acid residues in the binding cavity and found that the closest contacts with MG-TM are made by Ser89 and Ser91. The twist of azacyclopentane of MG-Pen was opposite to Ser89 and Ser91, indicating they have undergone greater repulsion. We further analyzed the conformation of N-substituents in MG-dL5** complex in detail (Fig. 3d and Fig. S4 in Supporting information). The nearest carbon atoms of MG-Pen to Ser89 and Ser91 are C22 and C9, which are 1.7 Å and 2.4 Å away from Ser91, respectively. The dihedral angle formed by C22 and C20-N19-C21 is 12.78°, and the dihedral angle formed by C9 and C8-N7-C11 is 40.61°, indicating that azocyclopentane underwent significant twisting due to the repulsion of Ser91. In MG-TM-dL5** complex, the shortest distance between the N-methyl group of MG-TM and the Ser91 is 3.1 Å and 3.6 Å, respectively. The dihedral angle formed by C1 and C8-N7-C9 is 0.14°, and the dihedral angle formed by C15 and C18-N17-C19 is 0.06°, indicating N-methyl group showed nearly no twisting conformation. Therefore, MG-TM would not be repelled in binding cavity, resulting in high binding affinity.
Although MG-Pen could enter the binding cavity of dL5** and wedge into the slot formed by side-chain of Tyr96, the binding affinity and stability are reduced dramatically due to the steric hindrance caused by the large-volume of azacyclopentane.
Subsequently, we assessed the binding affinity of MG-Pen and dL5** in practical applications in living cells. HeLa cells were transfected with pcDNA3.1-NLS-myc-dL5–2xG4S-mCer3 to co-express mCerulean3 (mCer3) fluorescent protein and dL5** in nuclei. We incubated these HeLa cells with 500 nmol/L MG-TM and MG-Pen respectively, then performed confocal imaging.
For the nuclei co-expressing mCer3 and dL5**, the 405-nm channel showed clear nuclear morphology with blue fluorescence from mCer3. MG-Pen and MG-TM binding to dL5** both showed enhanced fluorescence at 640-nm channel and overlapped well with the fluorescence of mCer3, indicating that the binding of MG to dL5** is highly specific (Fig. 3e). For HeLa cells stained with MG-TM, strong fluorescence was shown in 640-nm channel with a high signal-to-noise ratio (SNR) of 9-fold, while the cells incubated with MG-Pen, under the same imaging conditions, existed weak fluorescent intensity in 640-nm channel, and performed only 2-fold of SNR.
The decreased SNR of MG-Pen indicated that the weak binding affinity with dL5** led to the decline in binding stability, thereby resulting in the decline in binding density. This weak-binding imaging result is consistent with the results of molecular docking simulation.
PAINT technique relies on reversible and weak interaction between fluorescent probes and targets. Through capturing the fluorescence when probes instantaneous bind to the targets, and reconstructing the fluorescence signal accumulated from multiple imaging, PAINT could achieve single molecular localization and super-resolution imaging. Sparse labeling of targets is the basis of PAINT technology, to avoid the fluorescence signals from multiple targets within the diffraction limit overlapping in one capture, thereby achieving breakthroughs in the diffraction limit. Although the most representative DNA-PAINT have achieved extraordinarily high resolution, the DNA strand needs to be immunolabeled on the target, which limits the application in living cells. Nevertheless, PAINT imaging realized by MG-dl5** system could address this limitation effectively.
For PAINT imaging of dL5** conducted by traditional N-methyl MG-TM, in order to simultaneously meet the requirements of sparse labeling and dye replenishment under strong binding affinity, dye titration at extremely low concentrations (picomolar level) is required. However, the fine tune of the probe concentration resulted in long time-scale imaging (hour-level). We supposed that the weak binding affinity between MG-Pen and dL5** could satisfy low-density labeling under high-concentration of probes, which is conducive to continuous and rapid PAINT imaging.
We used MG-Pen to conduct dL5** PAINT imaging (Fig. 4a). We incubated HeLa cells expressing dL5** in nuclei with MG-Pen under different high concentrations (100 nmol/L, 500 nmol/L, and 1 µmol/L) for 30 min, respectively, and then performed PAINT imaging. We set the exposure time for each frame as 40 ms, and we acquired 3000 frames to complete one image, hence the imaging time for each image was only 2 min. The resolution of widefield was 453 nm, while the resolution of PAINT imaging reached 42.9 nm, showing the improvement of >10-fold (Figs. 4b–d).
Figure 4
Figure 4.
PAINT imaging of MG-Pen incubated with living HeLa cells expressing dL5** in nuclei. (a) PAINT imaging of nuclei under different concentrations of MG-Pen. Scale bar: 10 µm. Widefield (WF) imaging (b) and PAINT imaging (c) of dL5** in (a). (d) Intensity distributions across the white lines in (b) and (c). Localization accuracy (e) and average photons (f) of PAINT imaging under different concentration of MG-Pen (n = 3). Data are presented as mean ± standard deviation (SD). (g) PAINT imaging segmentally reconstructed in every 500 frames. Scale bar: 10 µm.
Under the concentration of 100 nmol/L of MG-Pen, PAINT imaging of the nucleus expressing dL5** showed a clear structure, and there was low background fluorescence outside the nucleus. The localization accuracy reached to 44.8 nm, and the average photon number was 377. As the concentration of MG-Pen increased to 500 nmol/L, the nucleus was able to show clear nuclear structure and dL5** localization under PAINT imaging, with a localization accuracy of 42.1 nm and an average photon number of 321. When the concentration of MG-Pen further increased to 1 µmol/L, we still obtained fine PAINT imaging of the nucleus, with a localization accuracy of 51.8 nm and an average photon number of 297 (Figs. 4e and f). Unfortunately, a significant background outside the nucleus was observed at 1 µmol/L of MG-Pen, due to the environmental sensitive and positively charged dye is easily to combine with various substances in cells (such as multiple membranes, proteins and nucleic acid).
As for the traditional N-methyl MG-TM, we performed PAINT imaging on HeLa cells expressing dL5** in the nucleus at the concentration of 500 nmol/L, and using the same imaging conditions as MG-Pen. Effective PAINT imaging could not be accomplished due to the strong binding affinity and high labeling density between MG-TM and dL5** (Fig. S5 in Supporting information).
To further evaluate the ability of MG-Pen to resist photobleaching and supplement labeling for continuous PAINT imaging at high concentrations, we segmentally reconstructed 3000 frames of PAINT imaging in every 500 frames, and obtained 6 reconstructed images at different imaging time-period (Fig. 4g). Under the concentration of 100 nmol/L of MG-Pen, only the image reconstructed from the first 500 frames have clear nuclear morphology. After 500 frames, there was no sufficient probes to supply the labeling of dL5**, resulting in insufficient fluorescence signal for reconstruction of images. As the concentration of MG-Pen increased to 500 nmol/L and 1 µmol/L, the initial 500 frames reconstruction presented clear nuclei, and as the imaging time increased, each subsequent 500 frames could also be effectively reconstructed to obtain PAINT images.
During PAINT imaging, when MG-Pen bound with dL5** was either dissociated or photobleached, the residual MG-Pen in the background solution continued to bind with dL5** as a supplement, thus continuously generating fluorescent signals at different targets to accomplish PAINT imaging. At the concentration of 100 nmol/L, a significant proportion of MG-Pen was consumed during the initial imaging period, resulting in insufficient residual MG-Pen and insufficient fluorescence for subsequent PAINT imaging. However, as the concentration of MG-Pen increased, although with substantial consumption of MG-Pen in the early period of imaging, there still remained an adequate amount of MG-Pen in the background solution to continually supplement the labeling of dL5**, thereby providing sufficient fluorescent signals throughout the entire PAINT imaging period.
In conclusion, we developed MG-Pen, a MG derivative with a bulky N-substituent, to enable rapid PAINT imaging of dL5**. By introducing a large azacyclopentane group, we significantly increased the steric hindrance of MG derivative binding with dL5**, which effectively reduced the binding affinity between MG-Pen and dL5**. This reduction in binding affinity allowed MG-Pen to maintain a low labeling density even at high concentrations, satisfying the sparse labeling requirements essential for PAINT imaging. Furthermore, the high concentration of MG-Pen in the background solution allowed for the sufficient replenishment of dissociated or bleached dyes, facilitating continuous imaging and thus enabling rapid PAINT imaging. In contrast to the traditional MG-TM, which required precisely dye titration and long-term imaging (several hours for an image), MG-Pen enabled the acquisition of a PAINT image in just 2 min with a localization accuracy of 42.1 nm. This significant improvement in imaging speed expands the potential of FAP-based systems for super-resolution microscopy, providing a powerful tool for real-time, high-resolution imaging of dynamic molecular processes in living cells. Moreover, the rapid PAINT imaging achieved with MG-Pen represents a crucial advancement in the field of live-cell imaging, promoting the development of time-resolved and dynamic super-resolution imaging techniques.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Xiangning Fang: Writing – original draft, Investigation, Data curation, Conceptualization. Qinglong Qiao: Supervision, Conceptualization. Fei Deng: Investigation. Zhaochao Xu: Writing – review & editing, Writing – original draft, Supervision, Conceptualization.
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Nos. 22225806, 22078314, 22278394, 22378385), Dalian Institute of Chemical Physics (Nos. DICPI202142, DICPI202436).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2025.110980.
[1]
Q. Qiao, W. Liu, J. Chen, et al., Angew. Chem. Int. Ed. 61 (2022) e202202961.
[2]
J. Chen, C. Wang, W. Liu, et al., Angew. Chem. Int. Ed. 60 (2021) 25104–25113. doi: 10.1002/anie.202111052
[3]
L. Miao, C. Yan, Y. Chen, et al., Cell Chem. Biol. 30 (2023) 248–260.
[4]
Q. Qiao, A. Song, K. An, et al., Angew. Chem. Int. Ed. 64 (2024) e202417469.
Figure 1
Mechanism of PAINT imaging and MG-dL5** imaging system. (a) Process and requirement of PAINT imaging. (b) Binding of MG and dL5** lead to fluorescence activation. (c) Molecule design of MGs with bulky N-substituents to achieve rapid PAINT imaging.
Figure 2
Photophysical properties of MGs with bulky N-substituents. (a–d) Ultraviolet–visible (UV–vis) spectra of MG-Aze (a), MG-Aze-F (b), MG-Aze-OH (c) and MG-Pen (d) in different solvents. (e) The fluctuation of normalized absorbance of MG-TM, MG-Aze, MG-Aze-F, MG-Aze-OH and MG-Pen in water during 1 h. (f) Mechanism of MG-Aze-F and MG-Aze-OH bleaching in water. The concentration of sample is 2 µmol/L.
Figure 3
Binding affinity evaluation of MGs. (a–c) Molecular docking analysis of MG-Aze (a), MG-Pen (b) and MG-TM (c). Left: frontal view of MGs/dL5** complex and calculated affinity; middle: spatial distribution of amino acid side chains in dL5** that contact MGs; right: electric charge density of MGs and binding cavity. (d) The twisting analysis of azacyclopentane of MG-Pen binding with dL5**. (e) Confocal imaging of Hela cells co-expressing mCer3 and dL5** in nuclei incubated with MGs (500 nmol/L). Scale bar: 10 µm.
Figure 4
PAINT imaging of MG-Pen incubated with living HeLa cells expressing dL5** in nuclei. (a) PAINT imaging of nuclei under different concentrations of MG-Pen. Scale bar: 10 µm. Widefield (WF) imaging (b) and PAINT imaging (c) of dL5** in (a). (d) Intensity distributions across the white lines in (b) and (c). Localization accuracy (e) and average photons (f) of PAINT imaging under different concentration of MG-Pen (n = 3). Data are presented as mean ± standard deviation (SD). (g) PAINT imaging segmentally reconstructed in every 500 frames. Scale bar: 10 µm.


 DownLoad:
DownLoad:



 下载:
下载:





