Table 1.
Representative marketed biomaterials for neural repair.
Citation:
Wenya Chi, Ruiyao Liu, Wenbo Zhou, Weilin Li, Yuan Yu. The mechanisms of interaction between biomaterials and cells/cellular microenvironment and the applications in neural injuries[J]. Chinese Chemical Letters,
2025, 36(8): 110587.
doi:
10.1016/j.cclet.2024.110587

The mechanisms of interaction between biomaterials and cells/cellular microenvironment and the applications in neural injuries
English
The mechanisms of interaction between biomaterials and cells/cellular microenvironment and the applications in neural injuries
Abstract:
Neural injuries can be induced by various neurological disorders and traumas, such as brain and spinal cord injuries, cerebrovascular diseases, and neurodegeneration. Due to the designable physicochemical properties, biomaterials are applied for various purposes in neural repair, including promoting axonal regeneration, reducing glial scar formation, delivering drugs, and providing temporary mechanical support to the injured tissue. They need to match the extracellular matrix (ECM) environment, support three-dimensional (3D) cell growth, repair the cellular microenvironment, mimic the tissue's biomechanical forces, and possess biodegradability and plasticity suitable for local intracavity applications. Meanwhile, functionalized biomaterials have been conducted to mimic the structural components of cellular ecological niches and the specific functions of the ECM. They can be engineered to carry a variety of bioactive components, such as stem cells and extracellular vesicles, which are used in neuroscience-related tissue engineering. Researchers also have developed biomaterial-based brain-like organs for high-throughput drug screening and pathological mechanistic studies. This review will discuss the interactions between biomaterials and cells, as well as the advances in neural injuries and engineered microtissues.
-
Key words:
- Biomaterial
- / Neural injury
- / Extracellular matrix
- / Bioscaffold
- / Nano-drug delivery system
- / Microtissue
-
1. Introduction
Neural injuries such as traumatic brain injury (TBI), peripheral neural injury, stroke, spinal cord injury (SCI), and neurodegenerative diseases (NDs) are significant causes of global mortality and disability. In the case of central nervous system (CNS) injuries such as TBI and SCI, shared disease progression involves primary and secondary injuries. The secondary injury involves a cascade of molecular, chemical, and inflammatory reactions that lead to further neural damage. Ischemic injury in stroke initiates a complex pathological cascade, resulting in excitotoxicity, mitochondrial dysfunction, neuronal loss, the release of excessive free radicals, protein misfolding, and inflammatory responses, ultimately culminating in nerve damage [1,2]. Peripheral neural injury is classified into five grades based on severity, with the most severe presentations involving neurotmesis, irreversible damage to axons, myelin sheaths, stroma, and potentially neurotoxins [3]. A defining pathological mechanism in NDs is protein denaturation, encompassing abnormal protein conformation and the deposition of proteins within neurons, culminating in neurological dysfunction [4]. Ultimately, diseases associated with neural damage have a microenvironment unfavorable for nerve regeneration, which can potentially lead to permanent impairment of neural function [5]. Current treatment approaches, including surgical and non-surgical methods, have limited effectiveness in achieving complete restoration of neural function.
The biomaterials have been widely studied for nerve repair by combining their in vivo biodegradation properties with programmed designs to make them suitable for the special physiological and pathological environments of neural injuries. They are available commercially for the repair of neural injuries, primarily to repair tissue defects and reconnect nerves (Table 1). However, these biomaterials have certain limitations such as insufficient adhesive strength, rapid degradation rates, and potential cellular toxicity. With the advancements in regenerative medicine and tissue engineering, new biomaterials are urgently required for clinical translation. This article focuses on the development of biomaterials in neuroscience, elucidates the mechanisms underlying the interaction between biomaterials and cells in their microenvironment, discusses the applications of biomaterials in neural injuries, and explores progress in engineering microtissues.
Table 1
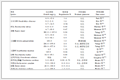
 DownLoad:
CSV
DownLoad:
CSV
Trade name Material Producer Main application Rayfilm Polylactic acid Medprin Regenerative Medical Technologies Co., Ltd. Absorbable dura mater patch Biodesign Small intestinal submucosal tissue from pigs (named SIS material) Cook Biotech Inc. Biological dural patch HaiAo Bovine skin tissue Yantai Zhenghai Bio-Tech Co., Ltd. Biofilm – Collagen extracted from bovine Achilles tendon TianXin Fu (Beijing) Medical Appliance Co., Ltd. Nerve Tape Porcine small intestine submucosal tissue BioCircuit Technologies, Inc. NeuroGen Animal collagen Integra LifeScience Nerve conduit NeuroFlex Animal collagen Regenity Biosciences AxoGuard Submucosa of porcine small intestine Axogen Tianyifu Animal collagen TianXin Fu (Beijing) Medical Appliance Co., Ltd. Avance Human collagen Axogen SaluTunnel Polyvinyl alcohol (PVA) Salunedica LLC Divine Bridge ECM Guangzhou Zhongda Medical Equipment Co., Ltd. Peripheral nerve defect repair materials Peripheral Nerve Sleeve Receiver Acetylated chitosan Beijing Huifukang Medical Technology Co., Ltd. For non-pathologic neural injuries of the median, ulnar, and radial nerves of the upper extremity (nerve defect length not greater than 2 cm) 2. Biomaterials and cells/cellular microenvironment
The cellular ecological niche comprises local tissue microenvironments that possess distinct biochemical, biophysical, and cellular properties, which help maintain cells in a specific state. Additionally, the molecular processes determining cell fate are diverse, with differentiation, proliferation, and other processes being crucial steps [6]. Biomaterials play a role in treating conditions such as TBI or NDs by participating in the regulation of the aforementioned molecular processes. Unaltered biomaterials influence specific cellular signaling pathways via inherent morphological characteristics or interactions with the cell nucleus, cytoplasm, and organelles. Biomaterials as carriers, control cells primarily through their bioactive components, such as stem cells, genes, and proteins, among other elements.
2.1 Cells and their microenvironment
Tissue repair and development involve the regulation of cell fate by intricate interactions between specific transcription factors and epigenetic networks in cellular niches. Components of the niche encompass soluble factors, such as growth factors, whereas interactions within the niche encompass cell-cell contacts and cell-matrix interactions [7]. The effect of the microenvironment on cell behavior varies among distinct cell types. For example, the microenvironment surrounding stem cells influences the quiescence, self-renewal, or differentiation of cells, whereas the microenvironment of somatic cells directs them to maintain specific cellular traits or initiate dedifferentiation or transdifferentiation.
Biochemical factors in the microenvironment mainly consist of soluble factors including growth factors and cytokines. These growth factors, secreted by stem cells or neighboring cells, serve as biochemical signals that maintain stem cell characteristics and regulate cell fate [8,9]. The function of growth factors is influenced by their concentration, and they can either diffuse away from the location where they are administered or become inactivated, thereby a controlled release method can be implemented using natural or synthetic materials to manage the distribution of growth factors and protect them from degradation within the body.
Biophysical signals within the microenvironment play an important role in controlling cell fate [10]. Biophysical signals encompass mechanical stimuli [11], direct communication between neighboring cells [12], compositions [13–16], morphology [17,18], and stiffness [19,20] of the extracellular matrix (ECM). The ECM constitutes a noncellular 3D macromolecular network, in which its components interact with cell adhesion receptors. Cell surface receptors are responsible for receiving signals from the ECM that help regulate a variety of cellular functions, including migration, immunity, and differentiation. The ECM continues to play a crucial role in regulating neural processes throughout development and adulthood, including synaptogenesis, axon growth, and synaptic stabilization [21]. Major ECM components include glycosaminoglycans (e.g., heparan sulfate and chondroitin sulfate), proteoglycans (such as neurocan and aggrecan), glycoproteins, and fibrin (collagen and fibronectin) [21]. Various ECM components fulfill distinct or overlapping functions. For instance, peptide sequences like Arg-Gly-Asp (RGD) and Pro-His-Ser-Arg-Asn within fibronectin can enhance cell adhesion to fibronectin [22]. The use of these sequences is frequently considered for incorporation into biomaterials to enhance cell-material adhesion. In both in vivo and in vitro settings, laminin acts as a crucial substrate for neural axon growth and has been integrated into scaffolds to improve cell adhesion and migration [23]. In the realm of biophysical signaling, substantial research has investigated the influence of ECM rigidity and structure on cellular behavior [24]. The stiffness of ECM affects various cellular processes, including migration, adhesion, spreading, differentiation, proliferation, and apoptosis [25,26]. The currently recognized mechanisms encompass cell recognition of changes in ECM stiffness through integrin or other cell surface adhesion molecules [27,28], involving signaling pathways, conformational alterations in mechanosensitive proteins (such as talin, vinculin, or lamin), and the activation of mechanosensitive ion channels [29,30]. Furthermore, cells respond to alterations in the nanomorphology of materials, which subsequently affect their migration, elongation, alignment, polarization, proliferation, and gene expression [31,32].
2.2 Biomaterial-mediated cell behavior
Functionalized biomaterials have been employed to guide the differentiation of stem cells into desired phenotypes or to modulate the dedifferentiation or transdifferentiation of somatic cells into cells of lower differentiation or distinct lineages [33]. Biomaterials predominantly influence cellular behavior by delivering chemical or bioactive molecules into target cells. The impact on cell differentiation is greatly influenced by the mechanisms of cellular uptake, including clathrin-mediated endocytosis, caveolin-mediated endocytosis, and caveolin-independent and clathrin-independent endocytosis [34,35]. During this process, intracellular mechanical stresses induced by biomaterials may significantly contribute to stem cell differentiation. Yi et al. observed that gold nanoparticles (Au-NPs) aggregated in perinuclear compartments and vesicular structures near the nucleus of mesenchymal stem cells (MSCs) [36]. This aggregation potentially led to interactions with proteins located in the cytoplasm, imparting mechanical stimuli to the MSCs. Protein kinase 38, a stress-activated protein kinase, was significantly upregulated in MSCs following interaction with Au-NPs, resulting in the enhanced osteogenic differentiation of MSCs.
Furthermore, the geometric shape of biomaterials plays a pivotal role in shaping cellular behaviors [37–39]. For instance, the aspect ratio of nanofibers and nanotubes (NTs) significantly influences cell fate regulation [40]. NTs with higher aspect ratios promote MSCs differentiation, whereas NTs with smaller aspect ratios fail to act as pro-differentiation agents. This diameter-dependent MSCs differentiation behavior is primarily attributed to the cytoskeletal tension and stress of MSCs grown on larger-diameter (70–100 nm) NTs [41,42]. On larger-diameter NTs, MSCs are compelled to elongate and stretch in their quest for protein clusters, leading to increased cytoskeletal tension and stress. This, in turn, enforces specific differentiation into osteocytes [42]. However, the majority of synthetic materials utilized for cellular microenvironments exist on a single scale, potentially missing the synergistic effects of other scales, which can result in inadequate structural integrity. To address this issue, Liu et al. developed a layered nanofiber cell matrix known as the "fiber-to-fiber" (FF) matrix, consisting of a network of gelatin nanofibers on ultra-thin fiber sheets [43]. The FF matrix combines the benefits of nanofibers and microfibers, including the promotion of cell growth and the enhancement of cell stability. Their findings demonstrated that the FF matrix effectively maintained pluripotency and normal chromosome morphology of human pluripotent stem cells (hPSCs) for over two months. Chen et al. utilized a microfluidic system to fabricate gelatin methacryloyl/chitosan (CS) microspheres incorporated into 3D bioprinted scaffolds [44]. The interaction between cells and the scaffold was investigated by co-culturing PC12 cells and Schwann cells. These findings suggest that the hydrogel multiscale composite scaffold with microspheres offers a favorable 3D microenvironment for the growth of neural synapses and could stimulate their growth and elongation. In addition, the biological orthogonal cross-linking or selective bioconjugation of biomaterials with cytokines, along with other techniques like click chemistry, cross-linking, modification, protein or cell targeting, and spatiotemporal material modification in the presence of living cells, has the potential to be synergistic in the treatment of neuroinflammatory diseases and other disorders [45]. In summary, by regulating the physicochemical properties of biomaterials, it is possible to create an environment conducive to cell adhesion, growth, and differentiation (Fig. 1) [46].
Figure 1
 Figure 1. Beneficial effects of biomaterials on cellular behavior and biology through the physical/chemical properties of biomaterials. Copied with permission [46]. Copyright 2018, The Author(s).
Figure 1. Beneficial effects of biomaterials on cellular behavior and biology through the physical/chemical properties of biomaterials. Copied with permission [46]. Copyright 2018, The Author(s).3. Biomaterials and neural injuries
After nerve damage, the chances for nerve regeneration diminish considerably owing to the chaotic microenvironment and the absence of structural support for nerve cells to anchor and form synapses. Natural and synthetic materials can be employed to repair damaged nerves, with common choices including poly(lactic-co-glycolic acid) (PLGA), gold, CS, and hyaluronic acid (HA). Biomaterials can be processed into various drug-loaded or non-drug-loaded forms such as NPs, hydrogels, and nanofibers. These materials can be used to repair damaged tissues or function as biological scaffolds, creating conducive microenvironments for neuronal cell regeneration. Moreover, these biomaterials can serve as delivery systems for stem cells, growth factors, exosomes (Exos), ribonucleic acid (RNA), and other agents aimed at promoting neural repair.
3.1 Post-injury ECM composition and biomaterials
In response to injury, the ECM undergoes alterations that vary depending on the specific site of the injury. Neurological disorders like Alzheimer's disease (AD) result from the gradual loss of nerve cells and alterations in ECM components in the brain [47]. Studies on AD have revealed that ECM components, including the restoration of 6-chondroitin sulfate levels in the perineuronal matrix network, display a protective effect on nerves that are tangled with tau protein [48]. Fibronectin, the major ECM protein, plays a crucial role in the growth of peripheral nerves and the regeneration of white matter [49]. One study shows that fibronectin treatment enhances serotonin axon sprouting [50]. In addition, the binding of fibronectin to cells, growth factors, and synthetic materials was found to promote axon growth [51].
In the spinal cord, the ECM can be diffusely distributed in the interstitial space or clusters around axons in Lang's nodes or synaptic junctions. In the context of SCI, certain components of the ECM can contribute to inflammation. For instance, HA fragments, matrix glycoproteins, tendon protein C, and fibronectin may function as endogenous Toll-like receptors (TLRs) ligands, triggering injury-associated molecular patterns and accelerating the inflammatory process [52]. When designing biomaterials, it is crucial to ensure their functionality in addressing or mitigating detrimental changes resulting from the mentioned injuries. For example, collagen has been shown to promote wound healing, cell adhesion, migration, proliferation, and functional nerve regeneration, and is widely used in various neurological injury disorders such as SCI, TBI, and peripheral nerve injury [53,54]. Silk fibroin has been shown to support cell attachment as well as nerve and Schwann cell proliferation, thereby promoting myelinated axon regeneration [5]. Fibrin has been shown to improve axon regeneration and support vascular repair, and can also promote cell adhesion, thereby improving nerve cell viability, proliferation, and differentiation [54,55]. Polyester has desirable mechanical properties, flexibility, porosity, etc., making it a valuable candidate for bioinks. However, evidence of material deformation has been observed under long-term strain. Many studies have shown that they support axonal regeneration, myelination, and the proliferation and differentiation of nerve cells [53,56,57].
Naturally, distinct types of neural injuries require specific biomaterials. Tubular materials are optimal for peripheral nerve regeneration and spinal cord axon repair, while gel-like materials are more suitable for brain repair. For instance, considering brain injuries, the composition of brain scars is notably softer than that of healthy tissue due to the absence of fibrous proteins like collagen [58]. Furthermore, the electromagnetic compatibility of brain structures imposes particular requirements for the utilization of biomaterials in the brain. First, these materials should avoid inducing inflammatory reactions. Second, biomaterials must exhibit minimal swelling to prevent an increase in intracranial pressure [59,60].
3.2 Biomaterial-based NPs for neural injuries
NPs are ultrafine particles with optical, electrical, and mechanical properties that can be controlled by various factors. These factors encompass hydrophilicity/hydrophobicity, surface charge, and particle size [61]. The selection of materials for the development of NPs is based on therapeutic requirements, considering factors such as their ability to evade the immune response, modification of targeting ligands, and the capacity to safeguard the effective drug payload in an injured microenvironment [62].
3.2.1 Nanoparticles and TBI
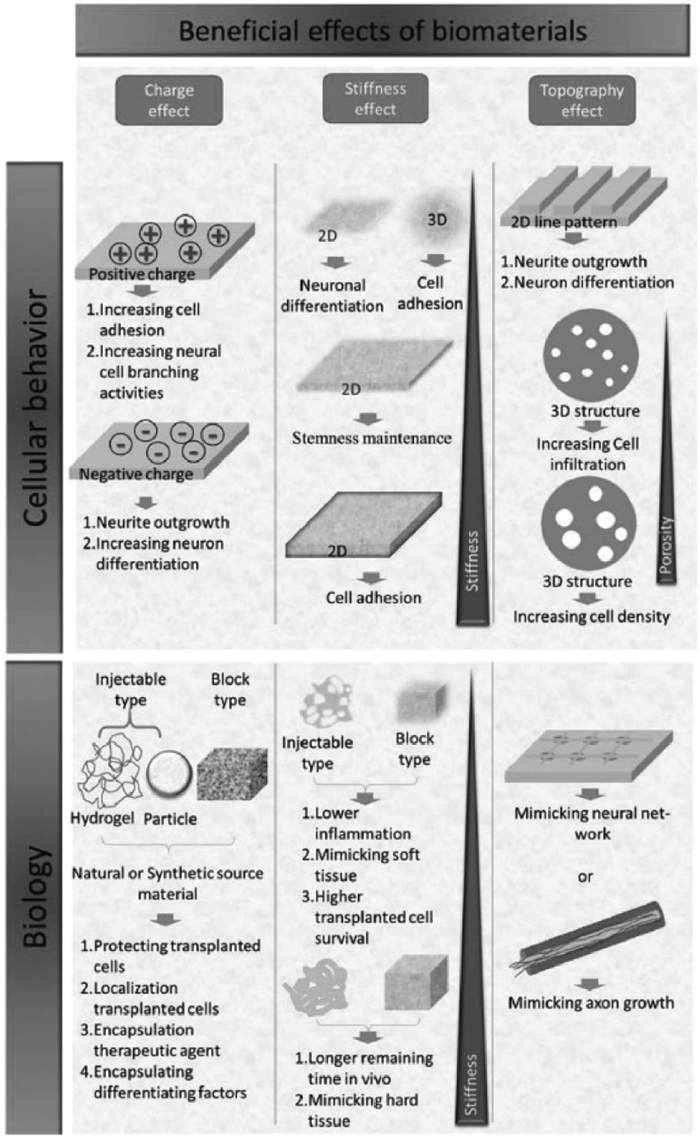
In the context of TBI, NPs can be customized for the potential to induce immunomodulation, support hemostasis, provide neuroprotection, act as antioxidants, and alleviate inflammation. NPs were functionalized with the synthetic peptide Gly-Arg-Gly-Asp-Ser (GRGDS) to promote hemostasis. This peptide, containing the RGD amino acid sequence, hinders platelet glycoprotein IIb/IIIa receptors, prevents fibrinogen binding, and regulates hemostasis [63]. Hubbard pioneered hemostatic nanoparticles (hDNPs) featuring surface modified GRGDS and encapsulated dexamethasone [64]. These hDNPs effectively reduce amygdala apoptosis, restore the compromised the blood-brain barrier (BBB), diminish astrocyte proliferation, and notably alleviate anxiety-like behavior. As shown in Fig. 2, Sun et al. formulated NPs, referred to as AMEC. These NPs were loaded with curcumin, functionalized with angiopep-2, and enriched with manganese [65]. When AMEC are administered intravenously, they efficiently cross the BBB through interactions with angiopoietin-2 and low-density lipoprotein receptor-associated protein-1. AMEC serves as scavengers for reactive oxygen species (ROS), reduces the ratio of macrophage M1/M2, and actively contributes to neuronal regeneration and the enhancement of functional recovery by combining the effects of eumelanin and curcumin.
Figure 2
 Figure 2. Bioactive multifunctional nanocomposite AMEC for targeted therapy and diagnosis of TBI with antioxidant and anti-neuroinflammatory effects. Copied with permission [65]. Copyright 2022, John Wiley & Sons, Inc.
Figure 2. Bioactive multifunctional nanocomposite AMEC for targeted therapy and diagnosis of TBI with antioxidant and anti-neuroinflammatory effects. Copied with permission [65]. Copyright 2022, John Wiley & Sons, Inc.3.2.2 NPs and spinal cord injury
Qi synthesized dextran NPs that were functionalized with ibuprofen and loaded with methylprednisolone (MP) through a direct esterification process involving the carboxylic acid of the hydrophobic drug and the hydroxyl group of the polymer backbone [66]. MP-loaded NPs reduced tumor necrosis factor-α (TNF-α) levels, significantly stimulated neurite growth, and reduced the degeneration of damaged neurons compared to intravenous MP alone. Since M1 macrophages exhibit an increased phenotype following SCI, it is common to develop NPs that promote the polarization of M1 macrophages towards the M2 phenotype. The anti-inflammatory agent etanercept (ET) was loaded onto a nanoflower structure made of molybdenum disulfide (MoS2) and poly(ethylene glycol) (PEG) [67]. ET-loaded MoS2@PEG exhibited the capacity to upregulate the production of anti-inflammatory factors, including interleukin-4 (IL-4) and IL-10, through the induction of M2-type macrophage polarization. Consequently, this resulted in the amelioration of inflammatory conditions following SCI and a remarkable enhancement in motor recovery in mice. It also evidenced that ET-MoS2@PEG nanomedicine could penetrate the injured spinal cord parenchyma within 96 h post-SCI, resulting in a noteworthy extension of the treatment timeline.
3.2.3 NPs and neurodegenerative diseases, stroke, and peripheral nerve injury
NPs have emerged as effective therapeutic diagnostic platforms for treating NDs such as AD, Parkinson's disease (PD), and epilepsy. Sun et al. developed polyethylene glycol-poly(lactic-co-glycolic acid) nanoparticles (bFGF-NPs) loaded with basic fibroblast growth factor (bFGF) using the emulsion/solvent evaporation method [68]. These were further conjugated with solanum tuberosum lectin (STL), which selectively binds to N-acetylglucosamine, facilitating nasal absorption and improving drug transport to the olfactory bulb, particularly to the brain. In addition, STL-bFGF-NPs enhanced choline acetyltransferase activity, which is neuroprotective for hippocampal neuronal cells of AD rats with AD. Novel NPs composed of CS and bovine serum albumin (BSA) have been designed to enhance drug penetration across the BBB. The anti-inflammatory agent curcumin is incorporated into these CS-BSA NPs, which stimulates microglial activation and enhances the removal of amyloid-β (Aβ) peptides through phagocytic activity [69]. Furthermore, curcumin-loaded CS-BSA NPs exhibited the ability to suppress the TLR4-mitogen-activated protein kinase (MAPK)/nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathway and downregulate M1 macrophage polarization. Mitochondrial dysfunction is a central pathogenesis of PD [70], Zheng et al. designed mitochondria-targeted nanoparticles (CSCCT NPs) that inhibited ROS production in 1-methyl-4-phenylpyridine-induced mitochondrial damage, and improved mitochondrial biogenesis by modulating the oxidized nicotinamide adenine dinucleotide (NAD)/silent information regulator 1 (SIRT1)/peroxisome proliferators-activated receptor γ coactivator lα (PGC-1α)/peroxisome proliferator-activated receptor γ (PPARγ)/nuclear respiratory factor 1 (NRF1)/mitochondrial transcription factor A (TFAM) pathway, which in turn improved mitochondrial dysfunction in neurons [71]. Ying et al. modified electrically responsive hydrogel nanoparticles (ERHNPs) with the brain-targeting peptide angiopep-2 (ANG) and loaded them with the antiepileptic drug phenytoin sodium (PHT) [72]. In the test model of amygdala ignition epilepsy, it was observed that ANG-ERHNPs could respond effectively to the abnormal brain discharges that occur during epileptic episodes by promptly releasing PHT.
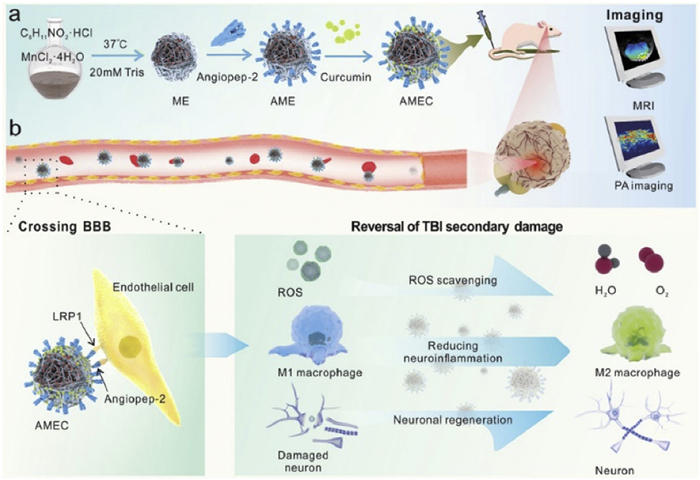
The biomimetic NPs have gained extensive usage in ischemic stroke owing to their notable features such as excellent biocompatibility, effective BBB penetration, and a strong inclination toward active homing. Neutrophil membrane-derived nanovesicles specifically target inflamed brain endothelial cells in a mouse model of ischemic stroke, delivering Resolvin D2 (RvD2) to the focal site and reducing the release of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, which protects the brain from perfusion injury [73]. Cell membrane-modified composite nanoparticles (CMNPs), when combined with neural stem cells (NSCs) that overexpress C-X-C chemokine receptor type 4 (CXCR4), have revealed the capacity to improve the efficacy of the NSCs, which were loaded with the anti-edema drug glibenclamide. This enhancement occurs through the targeting of CMNPs to the ischemic brain, facilitated by the interaction of CXCR4 with stromal cell-derived factor-1 (SDF-1) [74]. As shown in Fig. 3, Liu et al. designed erythrocyte (RBC) membrane-encapsulated brain-targeted bionic NPs loaded with curcumin, which acted by inhibiting the aberrant aggregation of α-synuclein (α-SYN), increasing the number of tyrosine hydroxylase (TH) neurons in the substantia nigra, providing antioxidant effects, and reversing mitochondrial dysfunction [75]. Table 2 [64–67,69,71–76] summarizes NPs for the treatment of neural injuries.
Figure 3
 Figure 3. RVG29-modified RBC membrane-encapsulated curcumin nanocrystals as novel biomimetic NPs with high BBB permeability properties and significant anti-PD effects. Copied with permission [75]. Copyright 2022, The Authors.
Figure 3. RVG29-modified RBC membrane-encapsulated curcumin nanocrystals as novel biomimetic NPs with high BBB permeability properties and significant anti-PD effects. Copied with permission [75]. Copyright 2022, The Authors.Table 2
Table 2. Nanoparticles for neural injuries.DownLoad:
CSV
Disease Administration Material Targeting receptor Modified ligand Loading agent Function and mechanism Ref. TBI Intravenous injection (Ⅳ) Polylactic acid-ethanolic acid-poly(ε-CBZ-L-lysine)-polyethylene glycol copolymer Platelet glycoprotein IIb/IIIa receptor GRGDS Dexamethasone Increased the number of microglia in the amygdala, decreased astrocyte proliferation, promoted repair of damaged BBB, and reduced anxiety-like behavior [64] Manganese Low-density lipoprotein receptor-related protein-1 (LRP-1) Angiopep-2 Curcumin Inhibited oxidative stress and neuroinflammation, promoted macrophage M1 phenotype polarization to M2 phenotype, and promoted neuronal regeneration [65] SCI Ⅳ Dextran based on ibuprofen modification \ \ MP Inhibited TNF-α, stimulated nerve growth and functional recovery [66] Molybdenum disulfide \ \ ET Macrophage polarized to an anti-inflammatory M2 phenotype, extravasated into injured spinal cord parenchyma, significantly prolonged treatment window [67] AD \ CS and BSA \ \ Curcumin Promoted phagocytosis of Aβ peptide by microglia, blocked the TLR4-MAPK/NF-κB signaling pathway, and reduced M1-type macrophage polarization [69] PD Ⅳ Cu, Se and 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-(polyethylene glycol)-triphenyl phosphate (DSPE-PEG2000-TPP) Neuronal mitochondria (vascular cell adhesion molecule-1 (VCAM-1)) Macrophage membranes (integrin) Curcumin Increased ratio of NAD to nicotinamide adenine dinucleotide (NADH) in neuronal mitochondria and increased expression of NAD-dependent deacetylase sirtuin-1 [71] Polyvinylpyrrolidone (PVP) and RBC membrane Acetylcholine receptors in BBB and neuronal cells Rabies virus peptide (RVG29) Curcumin Inhibited aberrant α-syn aggregation, promoted TH neuron expression, reversed mitochondrial dysfunction [75] Epilepsy Ⅳ 2-Dimethylamino ethyl methacrylate (DMAEMA) and N,N′-methylene bisacrylamide (MBA) LRP-1 Angiopoietin-2 PHT Shortened afterdischarge duration (ADD) and generalized seizure duration (GSD), reduced the post-discharge episodes [72] Apoplexy Ⅳ Neutrophil membrane Inflammatory endothelial cell neutrophil membrane RvD2 Targeted inflamed brain endothelial cells, reduced brain neutrophil infiltration, and relieved inflammation [73] Poly(lactic-ethanolic acid) SDF-1 Neural stem cell membrane (CXCR4) Glibenclamide Overexpression of CXCR4 enhanced NSCs migration to ischemic brain [74] Peripheral nerve injury Intramuscular injection (IM) CS Neuronal and tyrosine kinase receptor B Tetanus neurotoxin and BDNF Plasmid DNA that can encode BDNF Up-regulated the expression of BDNF, neurofilament heavy (NF-H), and growth associated protein-43 (GAP-43) in neural tissues, prevented long-term neurodegeneration and promoted nerve regeneration [76] 3.3 Biomaterials used as bioscaffolds for neural injuries
Bioscaffolds primarily employ biomaterials to provide substrates, both in vitro and in vivo, to promote cell anchoring, delivery, and growth. Bioscaffolds can be composed of various inorganic and organic materials. Alternatively, when high biocompatibility and tissue suitability are desired, scaffolds may incorporate natural materials that are components of the ECM, such as collagen, laminin, CS, HA, fibronectin, or decellularized matrices. These bioscaffolds facilitate reconstruction at the site of injury through mechanisms that include the release or production of active substances, attraction of native stem/progenitor cells to the scaffold site, modulation of the innate immune response, and specific induction of an anti-inflammatory macrophage phenotype.
3.3.1 Construction of bioscaffolds
Stem cells have garnered significant attention for their role in treating CNS diseases, particularly when they are incorporated into biological scaffolds. Stem cells possess the ability to self-renew and differentiate into various cell types, including embryonic stem cells, MSCs, induced pluripotent stem cells (iPSCs), and NSCs [77]. At the site of injury, stem cells can differentiate into the appropriate cell types, exerting therapeutic effects. Alternatively, they can promote neuroprotection, angiogenesis, and nerve regeneration through bystander effects including the secretion of neurotrophic factors [77]. Notably, the ECM of injured brain tissue cannot create a suitable environment for cell attachment, growth, and differentiation, rendering it unsuitable for the use of injectable stem cells [78,79]. For example, after brain injury in rats, bone marrow mesenchymal stem cells (BMSCs) were injected intravenously or intracardially, and only a minuscule fraction of the injected cells (<0.0005%) were observed at the injury site within 3 days [80]. In contrast, implantable bioscaffolds offer biomechanical support to cells, mimic the ECM, enhance cell survival, and localize stem cell aggregation and activity [79]. In a mouse stroke model, the implantation of scaffolds containing MSCs significantly improved cell viability compared to using MSCs alone [81]. Stem cells within these bioscaffolds can migrate to the damaged area, secrete neurotrophic factors, promote axonal regeneration and myelin formation, and reduce scar formation [82]. Mosiewicz et al. employed light-responsive scaffolds utilizing a light-controlled enzymatic formation. A caged peptide substrate was photoactivated and uncaged under ultraviolet (UV) light irradiation, allowing enzyme-catalyzed biomolecules to be locally tethered to the target region [83]. In addition, the scaffold could guide the migration of human mesenchymal stem cells (hMSCs). In another study, a microporous scaffold comprising chemically modified hydrogel microparticles effectively recruited neural progenitor cells to the stroke core after injection into the stroke region [84].
Exos, among the various types of extracellular vesicles, are small membrane vesicles secreted by eukaryotic cells that serve as essential mediators of intercellular communication and signaling [85]. Exos transport a wide range of biomolecules, including cytokines, proteins, lipids, chemokines, and nucleic acids, with the potential to profoundly influence cellular responses to injury and infection [86]. Furthermore, the membranes of Exos are equipped with various immune cell-related proteins, including major class I and class II histocompatibility complex proteins. These proteins play pivotal roles in antigen processing and presentation [87]. In a study conducted by Zhang et al., Exos from two sources was administered in rat models of brain injury. One group received 2D-cultured Exos, while the other group received Exos from MSCs cultured in 3D collagen scaffolds [88]. Both groups demonstrated the promotion of endogenous angiogenesis and neural regeneration. Notably, the group that received 3D-cultured MSC-Exos showed superior spatial learning outcomes. In another study, Exos secreted by human umbilical cord mesenchymal stem cells (hUMSCs) grown on 3D graphene scaffolds upregulated the expression of 68 miRNAs and significantly increased the expression of certain proteins, including neprilysin, insulin-degrading enzyme, and heat shock protein 70, in comparison to hUMSCs cultured on 2D graphene film [89].
3.3.2 Bioscaffolds and TBI
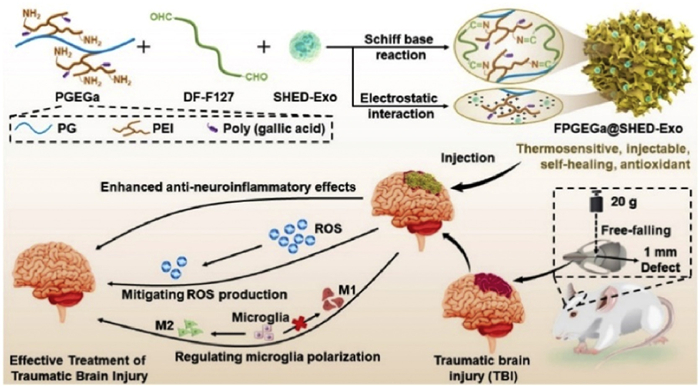
Biomaterials used as scaffolds can take different forms, such as fibers [90] and hydrogels [91]. Hydrogels are typically created through physical or chemical bonding or the self-organization of hydrophilic polymers. Their mechanical characteristics often resemble those of the brain matter. They can be formed by injecting them in liquid form, filling irregular injury cavities, and subsequently allowing them to polymerize [92]. Optimal bioscaffolds should be compatible with biochemical conditions (moisture and pH) and biophysical conditions (viscoelasticity and porosity) of the brain. These bioscaffolds should also possess the 3D structure of the ECM, offer physical support to the surrounding brain tissue, and act as a matrix for cell growth, axon regeneration, and neuronal protrusion formation [93]. Furthermore, they should promote cellular infiltration within bioscaffolds. Li et al. designed a bioactive antioxidant poly(citrate-gallate)-based hybrid hydrogel (FPGEGa) that encapsulated Exos derived from milky tooth pulp stem cells (SHED-Exos) for the treatment of TBI. As shown in Fig. 4, this hydrogel is thermosensitive, injectable, and antioxidant [94]. FPGEGa@SHED-Exos allowed an ultra-long sustained release of SHED-Exos for >21 days and significantly reduced intracellular ROS production in microglia. FPGEGa@SHED-Exos also improved neural regeneration in TBI rats by rescuing impaired motor function and regenerating damaged cortical tissue. Liu et al. fabricated scaffolds based on collagen and silk fibroin, incorporating Exos derived from hypoxia-preconditioned hUMSCs through 3D printing (3D-CS-HMExos) [95]. Hypoxia-preconditioned Exos are believed to possess pro-angiogenic factors. The results demonstrated that 3D-CS-HMExos doped with hypoxia-induced Exos significantly enhanced nerve regeneration and angiogenesis. Additionally, it suppressed neuronal apoptosis and the expression of pro-inflammatory factors, such as TNF-α and IL-6, while promoting the expression of the anti-inflammatory factor IL-10. Furthermore, the implantation of 3D-CS-HMExos offered an appropriate microenvironment for nerve regeneration, augmenting the development of axons and myelin at the site of injury.
Figure 4
 Figure 4. Engineered antioxidant poly(citrate-gallate)-Exo hydrogels with thermosensitizing, injectable, and self-healing capabilities modulate microglia immune phenotypes for TBI treatment. Copied with permission [94]. Copyright 2022, Elsevier Ltd.
Figure 4. Engineered antioxidant poly(citrate-gallate)-Exo hydrogels with thermosensitizing, injectable, and self-healing capabilities modulate microglia immune phenotypes for TBI treatment. Copied with permission [94]. Copyright 2022, Elsevier Ltd.3.3.3 Bioscaffolds and spinal cord injury
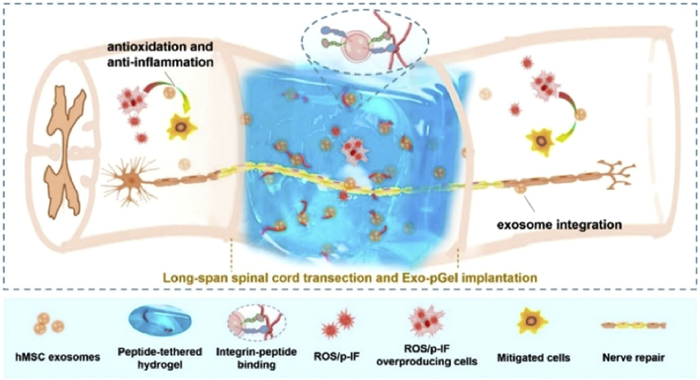
The similarities between spinal cord and brain injuries lie in the shared cascade of pathological events, encompassing initial cellular death and potential subsequent secondary injuries like neuroglial scar formation, inflammation, edema, ischemia, excitotoxicity, and heightened levels of free radicals [96]. As shown in Fig. 5, Li et al. established a novel implantation strategy using hMSCs-derived Exos and peptide-modified hydrogel (Exos-pGel) [23]. In contrast to systemic Exos delivery, localized implantation of Exos-pGel supplied damaged neural tissues with an ECM containing Exos. This approach successfully ameliorated the SCI microenvironment. The implanted Exos-pGel exhibited successful retention and continuous release within nerve tissue. To mimic the structure of a healthy spinal cord, researchers created a scaffold with collagen and silk proteins in a 3D configuration featuring hollow spaces [97]. This scaffold was then implanted with NSCs into laminectomized Sprague-Dawley (SD) rats. Rats in the 3D scaffold and NSCs groups displayed substantially higher functional neurological scores. MRI and diffusion tensor imaging confirmed ideal spinal cord continuity and injury cavity filling in the 3D scaffold and NSCs groups, respectively. Sun et al. utilized 3D printing to create a collagen-chitosan composite biomaterial scaffold, which was subsequently implanted into SD rats’ SCI models [98]. Their findings revealed that the 3D scaffold group exhibited the most significant regeneration of biotin-dextran amine profiles, reduced cavity formation, enhanced nerve fiber regeneration, and restoration of motor function. Thompson et al. developed HA hydrogels infused with ECM derived from astrocytes obtained from mouse embryonic stem cells [99]. This approach led to a reduction in glial scarring, promoted axon growth toward the lesion, and curtailed macrophage/microglia infiltration.
Figure 5
 Figure 5. Peptide-modified viscous HA hydrogel Exo-pGel containing hMSCs-derived Exos with antioxidant and anti-inflammatory effects promotes nerve repair after severe SCI. Copied with permission [23]. Copyright 2020, American Chemical Society.
Figure 5. Peptide-modified viscous HA hydrogel Exo-pGel containing hMSCs-derived Exos with antioxidant and anti-inflammatory effects promotes nerve repair after severe SCI. Copied with permission [23]. Copyright 2020, American Chemical Society.3.3.4 Bioscaffolds and neurodegenerative diseases, stroke, peripheral nerve injury
The mechanisms related to NDs, such as the misfolding of proteins, mitochondrial dysfunction, damage caused by oxidation, and responses associated with neuroinflammation, have been the subject of extensive research [100]. New research indicates that malfunctioning molecular chaperone-mediated autophagy (CMA) results in the buildup of harmful protein clumps in the CNS and plays a role in the development of NDs such as PD and AD [101]. Consequently, there is potential for safeguarding the CNS by enhancing CMA activity. Interestingly, in the context of acute neurological injuries, such as cerebral infarction, TBI, and SCI, it has been observed that CMA activity increases in the affected neural tissues. This finding suggests that augmenting the CMA pathway could also elicit neuroprotective effects in acute neural injuries.
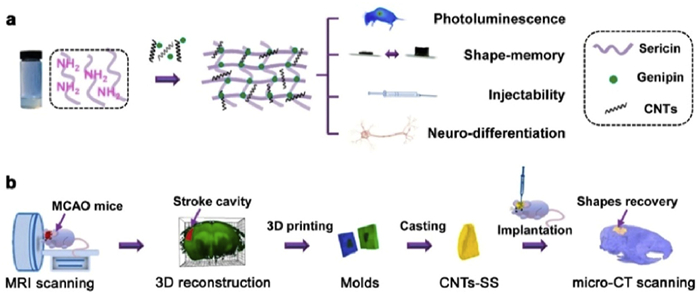
Wang et al. designed an injectable silk glycoprotein (SS) scaffold doped with photoluminescent carbon nanotubes (CNTs) [102]. As shown in Fig. 6, the natural cell adhesion properties of SS, combined with its unique neuroprotective activity, along with the pro-neural differentiation characteristics of CNTs, promote the differentiation of BMSCs into neurons. Additionally, the mathematically programmable memory properties of these scaffolds enable accurate filling of stroke cavity lesions. Ghuman et al. harnessed ECM to prepare hydrogels suitable for in situ injection into stroke cavities [103]. These ECM hydrogels not only attracted resident host cells but also activated the M2 phenotype of immune cells. Wang et al. utilized the HAMC hydrogel to locally release erythropoietin, which, in turn, attenuated the inflammatory response, reduced the size of the stroke cavity, stimulated the proliferation of endogenous neural stem and progenitor cells in the subventricular zone, and promoted brain repair after stroke injury in mice [104].
Figure 6
 Figure 6. An injectable, photoluminescent carbon nanotube-doped silk-gel scaffold CNTs-SS with programmable shape memory properties for individualized brain repair in severe ischemic stroke. Copied with permission [102]. Copyright 2021, The Authors.
Figure 6. An injectable, photoluminescent carbon nanotube-doped silk-gel scaffold CNTs-SS with programmable shape memory properties for individualized brain repair in severe ischemic stroke. Copied with permission [102]. Copyright 2021, The Authors.Liu et al. prepared a photocrosslinkable HA methacrylate hydrogel [105]. Soft hydrogels loaded with Exos offered superior healing for damaged peripheral nerves compared to hard hydrogels. Soft hydrogels rapidly release Exos, inhibiting macrophage infiltration and the secretion of pro-inflammatory factors IL-1β and TNF-α in damaged nerves. Chen et al. developed a nerve-guided catheter (NGC) loaded with the antioxidant melatonin (MLT) and magnetic iron tetraoxide nanoparticles (Fe3O4-MNPs) for sequential and sustainable drug release [106]. MLT scavenges free radicals, inhibits apoptosis by regulating neuroprotection-related enzymes and mitochondrial function, and exhibits antioxidant effects. Fe3O4-MNPs enhanced neuronal synapse growth by activating the mitogen-activated protein kinase signaling pathway. These composite scaffolds facilitate functional recovery, including myelin and axonal regeneration, similar to outcomes of autograft surgery in a model of sciatic nerve transection in treated SD rats [106]. In another study, NGC from polycaprolactone (PCL)/polyacrylic acid (PAA) composites was prepared by the electrohydrodynamic jet 3D printing process [107]. Compared to pure PCL scaffolds, PCL/PAA scaffolds exhibit a conductivity range similar to amphibian motor nerve fiber myelin sheaths, providing enhanced support for PC12 cell proliferation, growth, and neural differentiation. For relevant studies related to the delivery of bioactive components such as stem cells and Exos via bioscaffolds, please refer to Table 3 [23,94,95,108].
Table 3
Table 3. Bioscaffolds for neural injuries.DownLoad:
CSV
Disease Material Loading agent Function and mechanism Ref. TBI Collagen and silk protein Hypoxic preconditioning of hUMSCs-derived Exos Promoted nerve regeneration and angiogenesis, inhibited neuronal apoptosis, and promoted secretion of IL-10, enhanced recovery of locomotor function in TBI dogs [95] HA and collagen BMSCs-derived Exos Promoted the differentiation of NSCs into neurons and oligodendrocytes, promoted axon regeneration, myelin regeneration, synapse formation, and inhibited the differentiation of astrocytes [108] Poly(citrate-gallate)-based hybrid hydrogels Exos derived from human deciduous milk tooth pulp stem cells Reduced ROS production in microglia, promoted M2-type microglia polarization, and inhibited M1-type microglia polarization [94] SCI Aldehyde-modified HA and adipodihydrazide-modified HA and laminin-derived adhesion peptide PPFLMLLKGSTR (a motif in globular domain 3 of the human laminin-5 alpha3 chain) Human bone marrow mesenchymal stem cells derived Exos Reduced inflammation and oxidative damage, significantly promoted nerve recovery, and protected urinary tissues [23] 4. Engineered microtissues
Engineered microtissues refers to a biologically active tissue-like or organ-like structure created by co-printing biomaterials and cells using various techniques, such as 3D printing and microfluidics. Microtissues have valuable applications in regenerative medicine. In addition, they play a pivotal role in establishing bionic physiological and pathological models related to neurological injuries, which can be used for high-throughput drug screening and in-depth studies of pathological mechanisms.
4.1 Microtissue-engineered neural networks
Micro-TENNs, also known as microtissue-engineered neural networks, are 3D structures that replicate the neuroanatomy of the white matter pathways in the brain. These structures serve a dual purpose: they are designed as implantable microtissues for the reconstruction of axon tracts and for conducting anatomically relevant in vitro experiments [109].
Struzyna et al. developed micro-TENNs to promote neuronal survival and axonal extension [110]. These structures consist of discrete clusters of neurons integrated into axon bundles enclosed within miniature tubular hydrogels that contain ECM. Tubular hydrogels, measuring 180–500 µm in diameter and 2.0 cm in length, were fabricated by inserting longitudinally centered needles into cylindrical molds and filling them with liquid hydrogel. When injected into the rat brain using a stereotaxic apparatus, micro-TENNs led to Tenn neurons extending neural protrusions into the cortex. This resulted in the successful formation of synapses and preservation of extended axonal structures along the cortico-thalamic axis, which holds promise for restoring long-range axonal connections and rebuilding impaired brain pathways. Lai et al. integrated a pre-constructed functional neural network into the neural network of injured spinal cords [111]. This integration demonstrated the ability to promote the regeneration of functional axons and myelin through tropomyosin receptor kinase C (TrkC) signaling mediated by NT-3, ultimately leading to the restoration of functionality.
In a separate study, Sullivan et al. implanted micro-TENNs consisting of rat dorsal root ganglia and agarose hydrogel in a minimally invasive manner into the area of SCI [112]. Remarkably, axon growth originating from the graft was detected in the spinal cord, approximately 4.5 mm away from the site of implantation, five months after the injury occurred. Furthermore, micro-TENNs can be designed and optimized using computational models. Marinov et al. developed a new computational model based on a 3D diffusion equation that captures the unique features of 3D axon bundle development and simulates extensive unidirectional and bidirectional growth of actual neuronal structures [109]. Functions based on the concentration equation were employed to describe the growth, branching, rotation, and aggregation/bundling processes of each neuron, and these functions are distributed throughout the growth time of each growth segment.
4.2 3D bioprinting of brain-like organs
Organoids offer great promise for understanding intricate cell interactions within complex biological environments and hold immense potential in disease modeling and regenerative medicine [113]. By designing appropriate tissue microenvironments, organoids can exhibit spatially distinct organizational structures and morphologies [114]. In vitro brain-like organoids that resemble the brain are typically derived from IPSCs and cultured for several months in the presence of various growth factors and trophic support to drive organogenesis [113]. Matrigel R is the predominant biomaterial employed in brain-like organoid cultures. This enables the creation of intricate models that mimic specific human brain regions, including the formation of cortical plates [115], development of the forebrain [115], midbrain, and hypothalamus [116,117].
Restan Perez et al. utilized 3D printing to create hemispheres from adipose tissue-derived MSCs, employing fibrin-based bioinks to mimic brain tissue [118]. Subsequently, these constructs were subsequently incubated for eight days with a combination of SB431542, violet morpholino, LDN-193189, fibroblast growth factor 8, bFGF, and brain-derived neurotrophic factor (BDNF) to induce the differentiation of dopaminergic neurons (DN). Characterization of the neural tissue revealed high cellular activity, marked by the presence of early neuronal marker TUJ1 and DN marker TH, and the release of dopamine. Furthermore, upon exposure to potassium chloride, the neural tissue exhibited immature electrical signals. As this technology continues to mature, it is anticipated that neural tissues derived from MSCs will become valuable tools for modeling diseases and screening drugs related to PD. Lozano et al. employed a 3D printing system to construct brain-like structures consisting of distinct layers of neural cells encapsulated within gellan gum (GG) hydrogels modified with RGD peptides [119]. 3D-printed hydrogels were used to encapsulate primary cortical neurons and glial cells, resulting in increased cell survival and neural networks in RGD-coupled GG compared to single GG. Li et al. developed a 3D brain-like co-culture model in which star-shaped glial cells were embedded within a thermosensitive healing hydrogel [120]. Their approach utilized bioprinting techniques alongside NSCs to produce 3D neural spheroids.
4.3 3D bioprinting of nerve-like tissues
3D bioprinting involves the combination of cells with bioinks, which enable the fabrication of structures that closely resemble natural tissues. Neural-like tissues created through 3D bioprinting are primarily intended for in vitro neural tissue modeling. The bio-ink utilized in these preparations should closely resemble the ECM to enhance cell-cell interactions, proliferation, and differentiation [121]. Thus, Abelseth et al. developed an innovative bio-ink formulated with fibrin to facilitate the differentiation of human induced pluripotent stem cells (hiPSCs) into neural cells [55]. This bio-ink, comprising fibrinogen, goldfinch isoflavin, alginate, hiPSCs, CS, and thrombin, supports the derivation of hiPSCs into neural tissues, suggesting its potential for generating multiple engineered neural tissues. Joung et al. designed a 3D platform that mimics spinal cord tissue following a step-by-step process that involved the use of gelatin and fibronectin bioinks containing spinal neuronal progenitor cells (sNPCs) and oligodendrocyte progenitor cells (OPCs) derived from iPSCs [121]. This 3D scaffold incorporated multiple channels to facilitate the differentiation and axonal extension of sNPCs throughout the scaffold. Spontaneous calcium influx studies have confirmed the activity of these neuronal networks. Sharma et al. [122] employed a fibronectin-based bioink formulation combined with drug-loaded microspheres and NSCs derived from hiPSCs to print neural tissues. The microspheres contained small molecules, such as guggulsterone, which facilitated the differentiation of NSCs into DN. They used a microfluidic-based RX1 bioprinter, resulting in highly viable tissue with 95% cell survival seven days post-printing. This printed tissue expressed neural markers, including TuJ1, Forkhead box A2, TH, glial fibrillary acidic protein, and OPCs markers. qPCR analysis demonstrated the expression of TuJ1, NURR1, LMX1B, TH, and PAX6 in printed neural tissues after 30 days, indicating further development toward neurons. Gu et al. employed micro-extrusion-based bioprinting to create hydrogels encapsulating human neural stem cells (hNSCs) to generate 3D neural microtissues [123]. These hydrogels consist of alginate, agarose, and carboxymethyl cellulose. The encapsulated hNSCs successfully differentiated into neurons and formed functional synapses to establish the neural networks. Calcium imaging and scanning electron microscopy confirmed that these cells closely mimicked the morphology and activity of the human neural cells.
In summary, neural tissue structures are manufactured using multi-dispenser and coaxial extrusion bioprinting techniques. Ongoing research concentrates on enhancing cell survival, adhesion, and viability within these materials while preserving material printability. Furthermore, four-dimensional (4D) printing [124,125], an emerging method for creating biologics responsive to rapid external stimuli, addresses certain limitations inherent to 3D bioprinting technology, including the requirement for bioinks with precise viscosity [126].
5. Conclusions
The nervous system has a limited capacity for recovery due to the lack of inherent regeneration and repair mechanisms. The regenerative medicine and tissue engineering approaches offer promising strategies to enhance therapeutic outcomes. These methods utilize cells, biomaterials, and specific molecular or physical factors, either individually or in combination, to restore or replace damaged tissues, thereby promoting nerve repair and regeneration. The biological functions of biomaterials, particularly in regulating cell fate, have garnered increasing attention. Modifying biomaterials typically involves adjusting their chemical and physical properties to resemble natural tissues or incorporating growth factors, stem cells, and Exos into bioinks. These bioinks can create beneficial microenvironments that promote cell adhesion, proliferation, and differentiation. The development of biomaterials for different or similar pathomechanisms of nerve injury, such as inflammation, oxidative stress, and cell loss, can also be divided into different categories. For example, multifunctional nanocarrier systems, designed with targeting capabilities, can effectively cross the BBB and deliver therapeutic active molecules to specific sites of injury. Bioscaffolds are highly versatile in nerve injury repair due to their ECM-like structure, high porosity, and efficient transport of fluids and nutrients. Additionally, 3D printing technology enables the construction of micro-tissue-engineered neural networks, brain-like organoids, and neural-like tissues, which play a critical role in modeling neurological injuries. Regenerative medicine and tissue engineering have resulted in the development of various commercially available products for treating tissue injuries, including those affecting the skin, bone, and cartilage. However, due to the complex structural, physiological, and pathological characteristics of the nervous system, there are currently no comprehensive biomaterial-based products for neurological injuries on the market. As a result, further basic research is needed to clarify the safety, applicability, and efficacy of biomaterials in the treatment of neurological disorders.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Wenya Chi: Writing – original draft, Visualization. Ruiyao Liu: Writing – original draft. Wenbo Zhou: Writing – original draft. Weilin Li: Writing – original draft. Yuan Yu: Writing – review & editing, Funding acquisition, Conceptualization.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 82273487); the Young Medical Scientists Training Program (No. 21QNPY051); and the Shanghai Integration Achievement Program (No. 2022-RH17).
-
-
[1]
P.M. George, G.K. Steinberg, Neuron 87 (2015) 297–309.
-
[2]
W. Yu, E. Gong, B. Liu, et al., Chin. Chem. Lett. 34 (2023) 108205.
-
[3]
R. Sullivan, T. Dailey, K. Duncan, N. Abel, C.V. Borlongan, Int. J. Mol. Sci. 17 (2016) 2101. doi: 10.3390/ijms17122101
-
[4]
B.N. Dugger, D.W. Dickson, Cold Spring Harb. Perspect. Biol. 9 (2017) a028035.
-
[5]
M.E. Harley-Troxell, R. Steiner, R.C. Advincula, D.E. Anderson, M. Dhar, Polymers (Basel) 15 (2023) 3685. doi: 10.3390/polym15183685
-
[6]
H.V. Unadkat, M. Hulsman, K. Cornelissen, et al., Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 16565–16570. doi: 10.1073/pnas.1109861108
-
[7]
D.E. Discher, D.J. Mooney, P.W. Zandstra, Science 324 (2009) 1673–1677. doi: 10.1126/science.1171643
-
[8]
N. Tulina, E. Matunis, Science 294 (2001) 2546–2549.
-
[9]
L. Zhang, Q.S. Zuo, D. Li, et al., J. Integr. Agric. 14 (2015) 939–948.
-
[10]
L. Chen, Z. Yao, S. Zhang, et al., Chin. Chem. Lett. 34 (2023) 107925.
-
[11]
Y. Yang, K. Kulangara, R.T.S. Lam, R. Dharmawan, K.W. Leong, ACS Nano 6 (2012) 8591–8598. doi: 10.1021/nn301713d
-
[12]
C. Ottone, B. Krusche, A. Whitby, et al., Nat. Cell Biol. 16 (2014) 1045–1056. doi: 10.1038/ncb3045
-
[13]
D.S. Benoit, M.P. Schwartz, A.R. Durney, K.S. Anseth, Nat. Mater. 7 (2008) 816–823. doi: 10.1038/nmat2269
-
[14]
J.M. Curran, R. Chen, J.A. Hunt, Biomaterials 27 (2006) 4783–4793.
-
[15]
C.J. Flaim, S. Chien, S.N. Bhatia, Nat. Methods 2 (2005) 119–125. doi: 10.1038/nmeth736
-
[16]
Y. Mei, K. Saha, S.R. Bogatyrev, et al., Nat. Mater. 9 (2010) 768–778. doi: 10.1038/nmat2812
-
[17]
M.J. Dalby, N. Gadegaard, R.O. Oreffo, Nat. Mater. 13 (2014) 558–569. doi: 10.1038/nmat3980
-
[18]
K. Kulangara, Y. Yang, J. Yang, K.W. Leong, Biomaterials 33 (2012) 4998–5003.
-
[19]
D.E. Discher, P. Janmey, Y.L. Wang, Science 310 (2005) 1139–1143. doi: 10.1126/science.1116995
-
[20]
A.J. Engler, S. Sen, H.L. Sweeney, D.E. Discher, Cell 126 (2006) 677–689.
-
[21]
D. Lam, H.A. Enright, J. Cadena, et al., Sci. Rep. 9 (2019) 4159.
-
[22]
S. Aota, M. Nomizu, K.M. Yamada, J. Biol. Chem. 269 (1994) 24756–24761.
-
[23]
L. Li, Y. Zhang, J. Mu, et al., Nano Lett. 20 (2020) 4298–4305. doi: 10.1021/acs.nanolett.0c00929
-
[24]
M.P. Sousa, E. Arab-Tehrany, F. Cleymand, J.F. Mano, Small 15 (2019) 1901228.
-
[25]
M.C. Kim, Y.R. Silberberg, R. Abeyaratne, R.D. Kamm, H.H. Asada, Proc. Natl. Acad. Sci. U. S. A. 115 (2018) E390–E399.
-
[26]
V. Vogel, M. Sheetz, Nat. Rev. Mol. Cell Biol. 7 (2006) 265–275. doi: 10.1038/nrm1890
-
[27]
J.Z. Kechagia, J. Ivaska, P. Roca-Cusachs, Nat. Rev. Mol. Cell Biol. 20 (2019) 457–473. doi: 10.1038/s41580-019-0134-2
-
[28]
J.D. Humphrey, E.R. Dufresne, M.A. Schwartz, Nat. Rev. Mol. Cell Biol. 15 (2014) 802–812. doi: 10.1038/nrm3896
-
[29]
P.A. Janmey, D.A. Fletcher, C.A. Reinhart-King, Physiol. Rev. 100 (2020) 695–724. doi: 10.1152/physrev.00013.2019
-
[30]
O. Chaudhuri, J. Cooper-White, P.A. Janmey, D.J. Mooney, V.B. Shenoy, Nature 584 (2020) 535–546. doi: 10.1038/s41586-020-2612-2
-
[31]
R.G. Flemming, C.J. Murphy, G.A. Abrams, S.L. Goodman, P.F. Nealey, Biomaterials 20 (1999) 573–588.
-
[32]
H.J. Kong, D.J. Mooney, Nat. Rev. Drug Discov. 6 (2007) 455–463. doi: 10.1038/nrd2309
-
[33]
R. Olivares-Navarrete, S.L. Hyzy, D.A. Haithcock, et al., Bone 73 (2015) 208–216.
-
[34]
C. Brandenberger, C. Muhlfeld, Z. Ali, et al., Small 6 (2010) 1669–1678. doi: 10.1002/smll.201000528
-
[35]
H. Hillaireau, P. Couvreur, Cell Mol. Life Sci. 66 (2009) 2873–2896. doi: 10.1007/s00018-009-0053-z
-
[36]
C. Yi, D. Liu, C.C. Fong, J. Zhang, M. Yang, ACS Nano 4 (2010) 6439–6448. doi: 10.1021/nn101373r
-
[37]
B.A. Allo, D.O. Costa, S.J. Dixon, K. Mequanint, A.S. Rizkalla, J. Funct. Biomater. 3 (2012) 432–463. doi: 10.3390/jfb3020432
-
[38]
S. Zankovych, M. Diefenbeck, J. Bossert, et al., Acta. Biomater. 9 (2013) 4926–4934.
-
[39]
L. Zhang, T.J. Webster, Nano Today 4 (2009) 66–80.
-
[40]
K.D. Jandt, Evol. Adv. Eng. Mater. 9 (2007) 1035–1050. doi: 10.1002/adem.200700284
-
[41]
N. Wang, H. Li, W. Lü, et al., Biomaterials 32 (2011) 6900–6911.
-
[42]
S. Oh, K.S. Brammer, Y.S. Li, et al., Proc. Natl. Acad. Sci. U. S. A. 106 (2009) 2130–2135. doi: 10.1073/pnas.0813200106
-
[43]
L. Liu, K.I. Kamei, M. Yoshioka, et al., Biomaterials 124 (2017) 47–54.
-
[44]
J. Chen, D. Huang, L. Wang, et al., J. Colloid Interface Sci. 574 (2020) 162–173.
-
[45]
C. Cornelison, S. Fadel, Int. J. Mol. Sci. 23 (2022) 8496. doi: 10.3390/ijms23158496
-
[46]
Y.S. Chen, H.J. Harn, T.W. Chiou, Cell Transplant. 27 (2018) 407–422. doi: 10.1177/0963689717732991
-
[47]
S. Dauth, T. Grevesse, H. Pantazopoulos, et al., J. Comp. Neurol. 524 (2016) 1309–1336. doi: 10.1002/cne.23965
-
[48]
S. Yang, S. Gigout, A. Molinaro, et al., Mol. Psychiatr. 26 (2021) 5658–5668. doi: 10.1038/s41380-021-01208-9
-
[49]
V.J. Tom, C.M. Doller, A.T. Malouf, J. Silver, J. Neurosci. 24 (2004) 9282–9290.
-
[50]
C.Y. Lin, Y.S. Lee, V.W. Lin, J. Silver, J. Neurotraum. 29 (2012) 589–599. doi: 10.1089/neu.2011.2059
-
[51]
L.N. Novikova, L.N. Novikov, J.O. Kellerth, Curr. Opin. Neurol. 16 (2003) 711–715.
-
[52]
A.D. Gaudet, P.G. Popovich, Exp. Neurol. 258 (2014) 24–34.
-
[53]
T. Bedir, S. Ulag, C.B. Ustundag, O. Gunduz, Mater. Sci. Eng. C Mater. Biol. Appl. 110 (2020) 110741.
-
[54]
H.M. Khan, X. Liao, B.A. Sheikh, et al., J. Mater. Chem. B 10 (2022) 6859–6895. doi: 10.1039/d2tb01106a
-
[55]
E. Abelseth, L. Abelseth, L. De la Vega, et al., ACS Biomater. Sci. Eng. 5 (2019) 234–243. doi: 10.1021/acsbiomaterials.8b01235
-
[56]
M. Li, Y. Wang, J. Zhang, et al., J. Biomater. Sci. Polym. Ed. 29 (2018) 2168–2186. doi: 10.1080/09205063.2018.1528520
-
[57]
H. Amani, H. Kazerooni, H. Hassanpoor, A. Akbarzadeh, H. Pazoki-Toroudi, Artif. Cells Nanomed. Biotechnol. 47 (2019) 3524–3539. doi: 10.1080/21691401.2019.1639723
-
[58]
E. Moeendarbary, I.P. Weber, G.K. Sheridan, et al., Nat. Commun. 8 (2017) 14787.
-
[59]
A. Tuladhar, S.L. Payne, M.S. Shoichet, Front. Mater. 5 (2018) 14.
-
[60]
S. Mitragotri, J. Lahann, Nat. Mater. 8 (2009) 15–23. doi: 10.1038/nmat2344
-
[61]
S. Guo, Y. Liang, L. Liu, et al., J. Nanobiotechnol. 19 (2021) 32.
-
[62]
F.S. Mohammed, S.B. Omay, K.N. Sheth, J. Zhou, Expert. Opin. Drug Deliv. 20 (2023) 55–73. doi: 10.1080/17425247.2023.2152001
-
[63]
J.H. Beer, K.T. Springer, B.S. Coller, Blood 79 (1992) 117–128.
-
[64]
W.B. Hubbard, M. Lashof-Sullivan, S. Greenberg, et al., Sci. Rep. 8 (2018) 10622.
-
[65]
D. Sun, K. Liu, Y. Li, et al., Adv. Healthc. Mater. 11 (2022) e2200517.
-
[66]
L. Qi, H. Jiang, X. Cui, et al., Oncotarget 8 (2017) 99666–99680. doi: 10.18632/oncotarget.20649
-
[67]
G. Sun, S. Yang, H. Cai, et al., J. Colloid Interface Sci. 549 (2019) 50–62.
-
[68]
C. Zhang, J. Chen, C. Feng, et al., Int. J. Pharm. 461 (2014) 192–202. doi: 10.1007/978-3-662-45498-5_22
-
[69]
R. Yang, Y. Zheng, Q. Wang, L. Zhao, Nanoscale Res. Lett. 13 (2018) 330.
-
[70]
W. Poewe, K. Seppi, C.M. Tanner, et al., Nat. Rev. Dis. Primers 3 (2017) 17013.
-
[71]
Q. Zheng, H. Liu, H. Zhang, et al., Adv. Sci. 10 (2023) 2300758.
-
[72]
X. Ying, Y. Wang, J. Liang, et al., Angew. Chem. Int. Ed. 53 (2014) 12436–12440. doi: 10.1002/anie.201403846
-
[73]
X. Dong, J. Gao, C.Y. Zhang, et al., ACS Nano 13 (2019) 1272–1283.
-
[74]
J. Ma, S. Zhang, J. Liu, et al., Small 15 (2019) e1902011.
-
[75]
Y. Liu, J. Luo, Y. Liu, et al., ACS Cent. Sci. 8 (2022) 1336–1349.
-
[76]
C.D.F. Lopes, N.P. Goncalves, C.P. Gomes, M.J. Saraiva, A.P. Pego, Biomaterials 121 (2017) 83–96.
-
[77]
W. Zakrzewski, M. Dobrzyński, M. Szymonowicz, Z. Rybak, Stem Cell Res. Ther. 10 (2019) 68.
-
[78]
P.X. Wan, World J. Stem Cells 7 (2015) 448–460. doi: 10.4252/wjsc.v7.i2.448
-
[79]
L. Li, X. Chen, W.E. Wang, C. Zeng, Stem Cells Int. 2016 (2016) 9682757.
-
[80]
L.C. Turtzo, M.D. Budde, D.D. Dean, et al., PLoS One 10 (2015) e0126551. doi: 10.1371/journal.pone.0126551
-
[81]
L.N. Zamproni, M.A.V.M. Grinet, M.T.V.V. Mundim, et al., Nanomed. Nanotechnol. 15 (2019) 98–107.
-
[82]
M. Alvarado-Velez, S.F. Enam, N. Mehta, et al., Biomaterials 266 (2021) 120419.
-
[83]
K.A. Mosiewicz, L. Kolb, A.J. van der Vlies, et al., Nat. Mater. 12 (2013) 1072–1078. doi: 10.1038/nmat3766
-
[84]
K.L. Wilson, S.C.L. Pérez, M.M. Naffaa, S.H. Kelly, T. Segura, Adv. Mater. 34 (2022) e2201921.
-
[85]
B. Zhou, K. Xu, X. Zheng, et al., Signal Transduct. Target. Ther. 5 (2020) 144.
-
[86]
D.G. Phinney, M.F. Pittenger, Stem Cells 35 (2017) 851–858. doi: 10.1002/stem.2575
-
[87]
J. Kowal, G. Arras, M. Colombo, et al., Proc. Natl. Acad. Sci. U. S. A. 113 (2016) E968–E977.
-
[88]
Y. Zhang, M. Chopp, Z.G. Zhang, et al., Neurochem. Int. 111 (2017) 69–81.
-
[89]
L. Yang, Y. Zhai, Y. Hao, Z. Zhu, G. Cheng, Small 16 (2020) e1906273.
-
[90]
K. Nejati, D. Mehdi, S. Ghareghomi, et al., J. Drug Delivery Sci. Technol. 60 (2020) 102095.
-
[91]
M. Tsintou, K. Dalamagkas, T.L. Moore, et al., Neural Regen. Res. 16 (2021) 605–613.
-
[92]
M. Lacalle-Aurioles, C. Cassel de Camps, C.E. Zorca, L.K. Beitel, T.M. Durcan, Front. Cell. Neurosci. 14 (2020) 594304.
-
[93]
F.L. Maclean, M.K. Horne, R.J. Williams, D.R. Nisbet, APL Bioeng. 2 (2018) 021502.
-
[94]
Y. Li, M. Wang, M. Sun, et al., Compos. Part B: Eng. 242 (2022) 110034.
-
[95]
X. Liu, J. Wang, P. Wang, et al., Front. Bioeng. Biotechnol. 10 (2022) 1025138.
-
[96]
H. Nomura, C.H. Tator, M.S. Shoichet, J. Neurotraum. 23 (2006) 496–507. doi: 10.1089/neu.2006.23.496
-
[97]
J.P. Jiang, X.Y. Liu, F. Zhao, et al., Neural Regen. Res. 15 (2020) 959–968.
-
[98]
Y. Sun, C. Yang, X. Zhu, et al., J. Biomed. Mater. Res. A 107 (2019) 1898–1908. doi: 10.1002/jbm.a.36675
-
[99]
R.E. Thompson, J. Pardieck, L. Smith, et al., Biomaterials 162 (2018) 208–223.
-
[100]
L. Yan, M.-S. Guo, Y. Zhang, et al., Oxid. Med. Cell. Longev. 2022 (2022) 5288698.
-
[101]
H. Kanno, K. Handa, T. Murakami, T. Aizawa, H. Ozawa, Cells 11 (2022) 1205. doi: 10.3390/cells11071205
-
[102]
J. Wang, X. Li, Y. Song, et al., Bioact. Mater. 6 (2021) 1988–1999.
-
[103]
H. Ghuman, A.R. Massensini, J. Donnelly, et al., Biomaterials 91 (2016) 166–181.
-
[104]
Y. Wang, M.J. Cooke, C.M. Morshead, M.S. Shoichet, Biomaterials 33 (2012) 2681–2692.
-
[105]
Z. Liu, H. Tong, J. Li, et al., Front. Bioeng. Biotechnol. 10 (2022) 922570.
-
[106]
X. Chen, X. Ge, Y. Qian, et al., Adv. Funct. Mater. 30 (2020) 2004537.
-
[107]
S. Vijayavenkataraman, S. Thaharah, S. Zhang, W.F. Lu, J.Y.H. Fuh, Mater. Design 162 (2019) 171–184.
-
[108]
X. Liu, C. Wu, Y. Zhang, et al., Carbohyd. Polym. 306 (2023) 120578.
-
[109]
T. Marinov, H.A. Lopez Sanchez, L. Yuchi, et al., In Silico Biol. 14 (2020) 85–99. doi: 10.3233/isb-180172
-
[110]
L.A. Struzyna, J.A. Wolf, C.J. Mietus, et al., Tissue Eng. Part A 21 (2015) 2744–2756. doi: 10.1089/ten.tea.2014.0557
-
[111]
B.Q. Lai, M.T. Che, B.L. Du, et al., Biomaterials 109 (2016) 40–54.
-
[112]
P.Z. Sullivan, A. AlBayar, J.C. Burrell, et al., Tissue Eng. Part A 27 (2021) 1264–1274. doi: 10.1089/ten.tea.2020.0233
-
[113]
E. Di Lullo, A.R. Kriegstein, Nat. Rev. Neurosci. 18 (2017) 573–584. doi: 10.1038/nrn.2017.107
-
[114]
P. Hoang, Z. Ma, Acta Biomater. 132 (2021) 23–36.
-
[115]
M.A. Lancaster, N.S. Corsini, S. Wolfinger, et al., Nat. Biotechnol. 35 (2017) 659–666. doi: 10.1038/nbt.3906
-
[116]
J. Jo, Y. Xiao, A.X. Sun, et al., Cell Stem Cell 19 (2016) 248–257.
-
[117]
X. Qian, H.N. Nguyen, M.M. Song, et al., Cell 165 (2016) 1238–1254.
-
[118]
M. Restan Perez, R. Sharma, N.Z. Masri, S.M. Willerth, Biomolecules 11 (2021) 1250. doi: 10.3390/biom11081250
-
[119]
R. Lozano, L. Stevens, B.C. Thompson, et al., Biomaterials 67 (2015) 264–273.
-
[120]
Y.E. Li, Y.A. Jodat, R. Samanipour, et al., Biofabrication 13 (2021) 015014. doi: 10.1088/1758-5090/abc1be
-
[121]
D. Joung, V. Truong, C.C. Neitzke, et al., Adv. Funct. Mater. 28 (2018) 1801850.
-
[122]
R. Sharma, I.P.M. Smits, L. De La Vega, C. Lee, S.M. Willerth, Front. Bioeng. Biotechnol. 8 (2020) 57.
-
[123]
Q. Gu, E. Tomaskovic-Crook, R. Lozano, et al., Adv. Healthc. Mater. 5 (2016) 1429–1438. doi: 10.1002/adhm.201600095
-
[124]
M. Bodaghi, A.R. Damanpack, W.H. Liao, Smart Mater. Struct. 25 (2016) 105034. doi: 10.1088/0964-1726/25/10/105034
-
[125]
B. Liu, B. Dong, C. Xin, et al., Small 19 (2023) e2204630.
-
[126]
K. Osouli-Bostanabad, T. Masalehdan, R.M.I. Kapsa, et al., ACS Biomater. Sci. Eng. 8 (2022) 2764–2797. doi: 10.1021/acsbiomaterials.2c00094
-
[1]
-
Figure 1 Beneficial effects of biomaterials on cellular behavior and biology through the physical/chemical properties of biomaterials. Copied with permission [46]. Copyright 2018, The Author(s).
Figure 2 Bioactive multifunctional nanocomposite AMEC for targeted therapy and diagnosis of TBI with antioxidant and anti-neuroinflammatory effects. Copied with permission [65]. Copyright 2022, John Wiley & Sons, Inc.
Figure 3 RVG29-modified RBC membrane-encapsulated curcumin nanocrystals as novel biomimetic NPs with high BBB permeability properties and significant anti-PD effects. Copied with permission [75]. Copyright 2022, The Authors.
Figure 4 Engineered antioxidant poly(citrate-gallate)-Exo hydrogels with thermosensitizing, injectable, and self-healing capabilities modulate microglia immune phenotypes for TBI treatment. Copied with permission [94]. Copyright 2022, Elsevier Ltd.
Figure 5 Peptide-modified viscous HA hydrogel Exo-pGel containing hMSCs-derived Exos with antioxidant and anti-inflammatory effects promotes nerve repair after severe SCI. Copied with permission [23]. Copyright 2020, American Chemical Society.
Figure 6 An injectable, photoluminescent carbon nanotube-doped silk-gel scaffold CNTs-SS with programmable shape memory properties for individualized brain repair in severe ischemic stroke. Copied with permission [102]. Copyright 2021, The Authors.
Table 1. Representative marketed biomaterials for neural repair.
Trade name Material Producer Main application Rayfilm Polylactic acid Medprin Regenerative Medical Technologies Co., Ltd. Absorbable dura mater patch Biodesign Small intestinal submucosal tissue from pigs (named SIS material) Cook Biotech Inc. Biological dural patch HaiAo Bovine skin tissue Yantai Zhenghai Bio-Tech Co., Ltd. Biofilm – Collagen extracted from bovine Achilles tendon TianXin Fu (Beijing) Medical Appliance Co., Ltd. Nerve Tape Porcine small intestine submucosal tissue BioCircuit Technologies, Inc. NeuroGen Animal collagen Integra LifeScience Nerve conduit NeuroFlex Animal collagen Regenity Biosciences AxoGuard Submucosa of porcine small intestine Axogen Tianyifu Animal collagen TianXin Fu (Beijing) Medical Appliance Co., Ltd. Avance Human collagen Axogen SaluTunnel Polyvinyl alcohol (PVA) Salunedica LLC Divine Bridge ECM Guangzhou Zhongda Medical Equipment Co., Ltd. Peripheral nerve defect repair materials Peripheral Nerve Sleeve Receiver Acetylated chitosan Beijing Huifukang Medical Technology Co., Ltd. For non-pathologic neural injuries of the median, ulnar, and radial nerves of the upper extremity (nerve defect length not greater than 2 cm)  下载: 导出CSV
下载: 导出CSV
Table 2. Nanoparticles for neural injuries.
Disease Administration Material Targeting receptor Modified ligand Loading agent Function and mechanism Ref. TBI Intravenous injection (Ⅳ) Polylactic acid-ethanolic acid-poly(ε-CBZ-L-lysine)-polyethylene glycol copolymer Platelet glycoprotein IIb/IIIa receptor GRGDS Dexamethasone Increased the number of microglia in the amygdala, decreased astrocyte proliferation, promoted repair of damaged BBB, and reduced anxiety-like behavior [64] Manganese Low-density lipoprotein receptor-related protein-1 (LRP-1) Angiopep-2 Curcumin Inhibited oxidative stress and neuroinflammation, promoted macrophage M1 phenotype polarization to M2 phenotype, and promoted neuronal regeneration [65] SCI Ⅳ Dextran based on ibuprofen modification \ \ MP Inhibited TNF-α, stimulated nerve growth and functional recovery [66] Molybdenum disulfide \ \ ET Macrophage polarized to an anti-inflammatory M2 phenotype, extravasated into injured spinal cord parenchyma, significantly prolonged treatment window [67] AD \ CS and BSA \ \ Curcumin Promoted phagocytosis of Aβ peptide by microglia, blocked the TLR4-MAPK/NF-κB signaling pathway, and reduced M1-type macrophage polarization [69] PD Ⅳ Cu, Se and 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-(polyethylene glycol)-triphenyl phosphate (DSPE-PEG2000-TPP) Neuronal mitochondria (vascular cell adhesion molecule-1 (VCAM-1)) Macrophage membranes (integrin) Curcumin Increased ratio of NAD to nicotinamide adenine dinucleotide (NADH) in neuronal mitochondria and increased expression of NAD-dependent deacetylase sirtuin-1 [71] Polyvinylpyrrolidone (PVP) and RBC membrane Acetylcholine receptors in BBB and neuronal cells Rabies virus peptide (RVG29) Curcumin Inhibited aberrant α-syn aggregation, promoted TH neuron expression, reversed mitochondrial dysfunction [75] Epilepsy Ⅳ 2-Dimethylamino ethyl methacrylate (DMAEMA) and N,N′-methylene bisacrylamide (MBA) LRP-1 Angiopoietin-2 PHT Shortened afterdischarge duration (ADD) and generalized seizure duration (GSD), reduced the post-discharge episodes [72] Apoplexy Ⅳ Neutrophil membrane Inflammatory endothelial cell neutrophil membrane RvD2 Targeted inflamed brain endothelial cells, reduced brain neutrophil infiltration, and relieved inflammation [73] Poly(lactic-ethanolic acid) SDF-1 Neural stem cell membrane (CXCR4) Glibenclamide Overexpression of CXCR4 enhanced NSCs migration to ischemic brain [74] Peripheral nerve injury Intramuscular injection (IM) CS Neuronal and tyrosine kinase receptor B Tetanus neurotoxin and BDNF Plasmid DNA that can encode BDNF Up-regulated the expression of BDNF, neurofilament heavy (NF-H), and growth associated protein-43 (GAP-43) in neural tissues, prevented long-term neurodegeneration and promoted nerve regeneration [76]
下载: 导出CSV
Table 3. Bioscaffolds for neural injuries.
Disease Material Loading agent Function and mechanism Ref. TBI Collagen and silk protein Hypoxic preconditioning of hUMSCs-derived Exos Promoted nerve regeneration and angiogenesis, inhibited neuronal apoptosis, and promoted secretion of IL-10, enhanced recovery of locomotor function in TBI dogs [95] HA and collagen BMSCs-derived Exos Promoted the differentiation of NSCs into neurons and oligodendrocytes, promoted axon regeneration, myelin regeneration, synapse formation, and inhibited the differentiation of astrocytes [108] Poly(citrate-gallate)-based hybrid hydrogels Exos derived from human deciduous milk tooth pulp stem cells Reduced ROS production in microglia, promoted M2-type microglia polarization, and inhibited M1-type microglia polarization [94] SCI Aldehyde-modified HA and adipodihydrazide-modified HA and laminin-derived adhesion peptide PPFLMLLKGSTR (a motif in globular domain 3 of the human laminin-5 alpha3 chain) Human bone marrow mesenchymal stem cells derived Exos Reduced inflammation and oxidative damage, significantly promoted nerve recovery, and protected urinary tissues [23]
下载: 导出CSV
-







 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 0
- 文章访问数: 509
- HTML全文浏览量: 13
