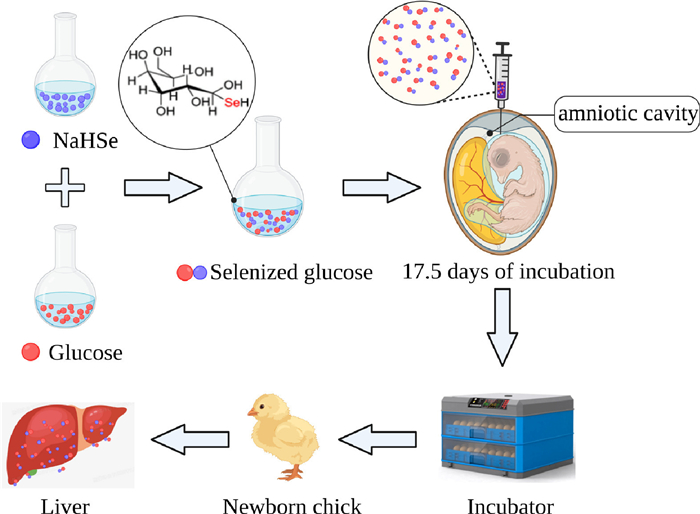
Figure 1.
Flowchart of in ovo feeding of selenized glucose and incubated in neonatal broilers.
The effects of in ovo feeding of selenized glucose on liver selenium concentration and antioxidant capacity in neonatal broilers
Jiahui Li , Qiao Shi , Ying Xue , Mingde Zheng , Long Liu , Tuoyu Geng , Daoqing Gong , Minmeng Zhao

The time for broilers to reach marketing age has been significantly shortened. It is consequently necessary to identify new nutritional solutions to improve the growth performance of broiler chickens [1]. The quality of chicks during the late embryonic and early hatching stages has an important impact on achieving superior production performance at marketing age. Unlike mammals, embryonic development of poultry is completely dependent on the nutritional reserves in the egg. During the latter stages of incubation, chicken embryos are exposed to a relatively hypoxic environment due to the increased demand for autogenous oxygen. At this time, the respiratory pattern changes from chorioretinal to pulmonary respiration. Therefore, the embryo is susceptible to oxidative stress during this process [2, 3]. Furthermore, oxidative stress can also be induced by the transition from a hypoxic to a fully oxygenated environment when newborn broilers emerge from the shell. This process may lead to a decrease in antioxidant capacity, growth retardation, and even increased mortality [4, 5]. In commercial production, newly hatched chicks usually do not have timely access to food and water for 24–72 h due to shelling time, management, and transportation factors [6]. Thus, post-hatch starvation may exacerbate existing oxidative stress and leading to a reduction in the antioxidant capacity and growth performance of broilers [7, 8].
Oxidative stress is closely associated with tissues of the liver, which is the main site of the production and consumption of reactive oxygen species (ROS). Inadequate regulation of oxidative stress increases the production of oxidative substances such as the superoxide (O2.−) and hydroxyl (OH.−), and hydrogen peroxide (H2O2). These reactive oxygen species can inhibit the activity of antioxidant enzymes and cause oxidative damage in liver tissue [9]. Consequently, it is important to enhance the hepatic antioxidant capacity of late embryos and neonatal broilers.
In ovo feeding (IOF) is a technique whereby different exogenous nutrients are injected into eggs via the amniotic cavity. It is typically performed on the day 17th or 18th of incubation. The principle is that the amniotic fluid can be swallowed by the embryo during the later stages of incubation. Hence, the injection solutions containing supplemental nutrients can be directly absorbed and utilized by the embryo, thereby enhancing the growth and development of poultry. Numerous researchers have reported that IOF of exogenous nutrients such as carbohydrates, amino acids, and their metabolites can increase hatching weight, liver glycogen reserves, marketing weight, and breast muscle yield [10, 11]. In addition, supplementing vitamin E in eggs can enhance the antioxidant capacity of chicks [12]. Consequently, using IOF technology could ameliorate the oxidative stress of late embryonic and neonatal broilers.
Selenium (Se) is a non-metal trace element that plays essential roles in maintaining the normal physiological functions of the liver [13]. Se typically occurs as the active center of selenoproteins [14]. Se can enhance the activity of selenoproteins such as glutathione peroxidase (GSH-Px) and thioredoxin reductase (TrxR) [15]. Se also plays physiological roles as a scavenger of free radicals, thereby preventing lipid peroxidation and promoting antioxidant activity [16]. In nature, Se generally exists in the form of either inorganic or organic Se compounds, among which, the former mainly include sodium selenite and sodium selenate selenium sulfide, which are characterized by high toxicity, poor absorption, and low bioavailability. Contrastingly, organic Se compounds, such as Se yeast (SY) and seleno-substituted amino acids, are demonstrated to have minimal toxicity, efficient absorption, and high bioavailability. However, the excessive costs limit the application of organic Se in livestock production [17]. Selenized glucose (SeGlu) is a newly invented organoselenium material [18] with high chemical stability and low toxicity [19]. It has already been widely applied in producing the Se-enriched agricultural products [20], which are popular nowadays because selenium is a key element for the health of human nervous, cardiovascular and hepatobiliary systems [21, 22]. Studies have shown that dietary supplementation with SeGlu can enhance the antioxidant activity of egg yolk and liver antioxidant capacity in laying hens [23, 24]. On the basis of these observations, we hypothesized that IOF of SeGlu would enhance Se reserves and antioxidant ability in newborn chicks. In this study, we thus sought to confirm this supposition by evaluating the influence of IOF of SeGlu on the antioxidant capacity of liver in neonatal broilers (Fig. 1).
Compared with the control group (0.75% physiological saline), we found that the SeGlu10 (IOF of 10 µg Se from SeGlu) and SeGlu20 (IOF of 20 µg Se from SeGlu) treatments promoted significant increases in the Se content of broiler livers (Table 1). In response to an increase in SeGlu concentration, we detected a linear change pattern in the Se contents of livers in newborn broilers. However, there were no significant differences among the three treatment groups with respect to the hatching weight of broiler chicks.
 DownLoad:
CSV
DownLoad:
CSV

|
As shown in Table 2, compared with control group birds, broilers in the SeGlu20 group were characterized by significant increases in GSH-Px activity. Polynomial orthogonal contrast analysis revealed that the effects of SeGlu concentration on GSH-Px activity were both linear and quadratic, and that the latter model provided a better fit to the data. Compared with the control group, IOF of 10 µg Se was found to promote a significant reduction in liver glutathione (GSH) concentration, and we established that the response of GSH levels to an increase in SeGlu concentration was quadratic. Contrastingly, we detected no appreciable differences among treatment groups on glutathione reductase (GR) and TrxR activity.
DownLoad:
CSV

|
The effects of in ovo SeGlu administration on the free radical scavenging capacity of livers in newborn broilers are shown in Table 3. In response to an increase in SeGlu levels, we detected a linear increase in liver total superoxide dismutase (T-SOD) activity in newly hatched chicks, whereas in contrast, there was no clear difference among the three treatments on liver total antioxidant capacity (T-AOC) activity. When administered at a concentration of 10 µg, Se was observed to promote a significant increase in the scavenging of O2.− in liver compared with that detected in the other two groups, whereas we observed no significant difference among the three groups with respect to OH.− scavenging ability. Additionally, compared with control group broilers, those in the SeGlu10 group were found to have lower liver concentrations of malondialdehyde (MDA), and we established that MDA levels responded to SeGlu dosage in significant quadratic manner.
DownLoad:
CSV

|
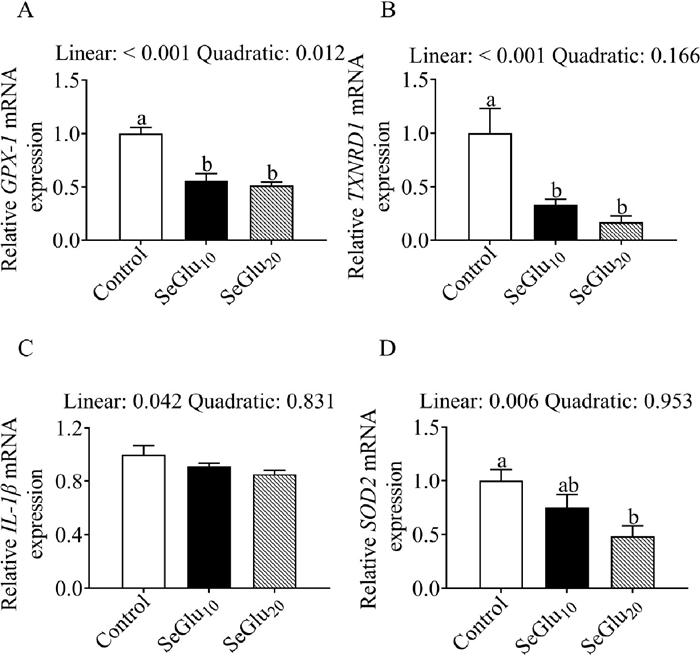
As shown in Fig. 2, IOF of SeGlu decreased the mRNA expression of GPX-1, TXNRD1, and SOD2 mRNAs, whereas these treatments appeared to have no appreciable effect on IL-1β transcript levels.
Previous studies have established that dietary supplementation with Se can increase the Se content in tissues and eggs, thereby indicating that supplementary feeding can enrich Se levels in animals [25, 26]. Although Se is an essential trace nutrient in poultry production, chicken embryos do not have access to exogenous Se during the period of incubation [27]. Se concentrations in fertilized eggs are essentially determined by its level in the diet of laying hens [28]. Consequently, supplementation of maternal feed with Se is the traditional approach employed to enhance Se storage in embryonic chicks. However, Ohta et al. found that in ovo administration of amino acids can contribute to enhancing the corresponding levels of amino acids in broiler embryos [29]. Similarly, Saeed et al. demonstrated that the injection of 80 or 120 µg folic acid elevated folic acid concentrations in the blood of broilers measured on day 42 post-hatch [30]. Furthermore, several researchers indicated that injecting of carbohydrates, including sucrose and maltose, can enhance the glycogen reserves of domestic pigeons and broilers [31, 32]. These findings imply that IOF of exogenous nutrients into avian eggs could serve as an alternative approach to enhancing the nutritional status of embryos and hatchlings. In line with expectation, we established that IOF of SeGlu promoted a linear increase in Se contents in the livers of newborn broilers. Similarly, in our previous studies, we found that SeGlu supplementation contributed to elevating Se levels in the eggs, liver, oviduct, and spleen of laying hens [23, 24].
Se deficiency has been established to adversely affect the expression of selenoproteins in immune-related organs, thereby inhibiting the pleiotropic regulatory mechanisms of organisms in response to oxidative stress [33]. However, excessive intake of Se may also cause toxicity, which can lead to severe immunosuppressive effects and disease in animals [34]. In the present study, we found that injection of 10 or 20 µg Se from SeGlu had no significant effect on either the hatchability (data not shown) or weight of newborn broilers. These results are consistent with those reported by Lee et al. [35], who found that IOF of Se at concentrations ranging from 0.075 µg to 20 µg per egg had no evident influence on the hatchability or body weight of broilers on the day of hatching. These findings indicate that the dosages of SeGlu used in the present study are safe for the nutrient supplementation of broiler embryos. It is well established that the physiological effects of Se in animals are mediated primarily via the activity of this element as a functional component of selenoproteins [14]. The major sources of organic Se additive in poultry production are Se-containing amino acids, such as selenomethionine, selenocystine, and selenocysteine [18]. Our observations indicate that glucose can also be used as the carrier of elemental Se. The reason may be the catabolic intermediates of glucose can serve as the carbon skeletons for synthesizing amino acids. Accordingly, IOF of SeGlu can be considered a further effective approach for enhancing Se reserves in the livers of neonatal broilers.
In addition to being the main site for regulating the effects of oxidative stress, the liver also carries out the synthetic metabolism of Se to selenoproteins [36]. The endogenous antioxidant system of animals is dependent on the selenoprotein system, which consists mainly of enzymes in the GSH and TrxR families [27, 37]. In particular, GSH-Px and TrxR can assist in catalyzing the reduction of H2O2 and organic peroxides. TrxR uses nicotinamide adenine dinucleotide phosphate to maintain the reduced state of thioredoxin, and also plays roles in scavenging ROS, thereby protecting cells from the oxidative damage caused by free radicals [38]. It has previously been shown that IOF of nano-selenium produced by lactic acid bacteria can significantly increase the activities of antioxidant enzymes such as catalase, SOD, and GSH-Px in chicks [39]. Similarly, IOF of sodium selenite, selenium yeast or selenomethionine has been shown to be beneficial in enhancing the antioxidant capacity and immune function of chicks [40]. In the present study, we found that IOF of SeGlu promoted an increase in the GSH-Px activity of broiler livers, and similar effects have been reported in laying hens fed SeGlu-supplemented diets [23, 41]. Moreover, Yuan et al. have found that selenium yeast can significantly enhance GSH-Px mRNA expression in broiler livers [42]. Xiao et al. found that dietary supplementation with Se enhances GSH-Px and TrxR activities and the levels of GPX1 and TXNRD1 mRNA expression [43]. Interestingly, in the current study, IOF of SeGlu increased GSH-Px activity, but resulted in a decrease in GPX1, TXNRD1 mRNA expression in the liver of broilers. Moreover, previous researchers indicated that the activities of GR and TrxR, as well as T-AOC in chicken were significantly elevated after Se supplementation in the diet [44, 45]. However, neither the activities of GR and TrxR nor T-AOC in liver was altered by SeGlu injection. We suspect that these discrepant observations could be associated with the breed of test chicken, length of experimental observations, method of administration, or may indeed be attributable to differences in metabolic pathways of the administered form of Se. Consequently, we speculate that SeGlu administered via the in ovo route may primarily enhance the antioxidant capacity of chicks by elevating the enzymatic activity of the GSH system.
The amount of MDA is often negatively correlated with the GSH-Px activity and can reflect the antioxidant status of the body [46]. We found that IOF of 10 µg Se reduced the MDA contents in the liver of broiler chicks. This is consistent with the findings of Wang and Xu [47], who demonstrated that Se enhances antioxidant activities in the serum and liver of animals. These findings provide evidence to indicate that in ovo administration of SeGlu enhances the efficacy of the liver's antioxidant defense system. It is well established that the three endogenous reactive oxygen species O2.−, OH.−, and H2O2 are the main sources of ROS-associated oxidative stress in cells. Excessive ROS production induces DNA damage, apoptosis, and inflammation [48, 49]. Therefore, the antioxidant mechanism of scavenging endogenous ROS is essential for the health of the organism, and the FRSC of oxygen radical in the organism can reflect the antioxidant capacity of the animal [50]. In the present study, we detected elevated levels of O2.− scavenging in the livers of chicks that had been subjected to the SeGlu10 treatment, which is consistent with the observation of Chen et al. [51], who detected an enhancement of free radical scavenging capacity in response to the administration of Se. Therefore, IOF of SeGlu can enhance the antioxidant capacity of chick livers and also reduce the sensitivity to tissue peroxidation.
As an important natural antioxidant enzyme, SOD plays key roles in the association between oxidation and antioxidant balance in animals. SOD acts as the first line of the antioxidant defense system by promoting the degradation of superoxide anions [52]. Of the three SOD isoforms found in animals, SOD2 occurs mainly in the mitochondrial matrix [53]. ROS are primarily released into the mitochondrial matrix, so SOD activity and SOD2 expression are generally closely associated with cellular antioxidant capacity. Guido et al. [43] found that dietary supplementation with organic Se can contribute to a significant increase in SOD activity in the livers of laying hens, which is similar to the findings of Xiao et al. [54]. In the current study, the chicks in SeGlu treatment had relatively high T-SOD activity but reduced SOD2 mRNA expression. This suggests that IOF of SeGlu can increase T-SOD activity to scavenge ROS. Hence, IOF of SeGlu may increase the antioxidant ability of liver through enhancing the T-SOD activity in broilers.
SeGlu is a newly invented organoselenium material and is widely used. However, its mechanism was not deeply studied in previous works. This paper provided an in-depth study on the mechanism of SeGlu to enhance the antioxidant capacity of liver of newborn chicks using in ovo technology. In summary, IOF of SeGlu can promote increases in selenium contents in the livers of neonatal broilers, and can also contribute to enhancing the antioxidant capacity and decreasing the MDA level by elevating the activities of antioxidant enzymes such as T-SOD and GSH-Px. These findings provide a basis for subsequent work on the study of Se-enriched additive in agricultural animal.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported by the Open Project of Jiangsu Key Laboratory of Animal genetic Breeding and Molecular Design (No. AGBMD202202); the Jiangsu Agricultural Science and Technology Innovation Fund (Nos. [CX(21)3131] and [CX(20)3010]); the "JBGS" Project of Seed Industry Revitalization in Jiangsu Province (Nos. [JBGS[2021]027 and JBGS[2021]105]); the Science and Education Integration Project of Yangzhou University (No. [KJRH202115]); and Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Supplementary material associated with this article can be found, in the online version, at doi:
C. Ricks, N. Mendu, P. Phelps, Poult. Sci. 82 (2003) 931–938. doi: 10.1093/ps/82.6.931
E. Jassim, M. Grossman, W. Koops, et al., Poult. Sci. 75 (1996) 464–471. doi: 10.3382/ps.0750464
S. Giusti, D. Converso, J. Poderoso, et al., Eur. J. Neurosci. 27 (2008) 123–131. doi: 10.1111/j.1460-9568.2007.05995.x
M. Stock, K. Silvernail, J. Metcalfe, Free Radic. Biol. Med. 8 (1990) 313–318. doi: 10.1016/0891-5849(90)90095-Z
Z. Uni, L. Yadgary, R. Yair, J. Appl. Poult. Res. 21 (2012) 175–184. doi: 10.3382/japr.2011-00478
Y. Noy, Z. Uni, World's. Poult. Sci. J. 66 (2010) 639–646. doi: 10.1017/S0043933910000620
J. Ge, H. Li, F. Sun, et al., J. Anim. Sci. 9 (2017) 2871–2878.
F. Sun, Y. Zuo, J. Ge, et al., Poult. Sci. 97 (2018) 2638–2646. doi: 10.3382/ps/pey146
M. Farzaei, M. Zobeiri, F. Parvizi, et al., Nutrients 10 (2018) 855. doi: 10.3390/nu10070855
W. Zhai, D. Rowe, E. Peebles, Poult. Sci. 90 (2011) 1295–1301. doi: 10.3382/ps.2010-01130
S. Tadeusz, P. Jan, G. Stanisław, et al., BMC Vet. Res. 15 (2019) 105. doi: 10.1186/s12917-019-1850-8
M. Karageçili, E. Babacanoğlu, Br. Poult. Sci. 18 (2022) 1–7.
M. Michalczuk, M. Batorska, U. Sikorska, et al., Anim. Sci. J. 92 (2021) e13662. doi: 10.1111/asj.13662
M. Rayman, Lancet 379 (2012) 1256–1268. doi: 10.1016/S0140-6736(11)61452-9
S. Holger, S. Bodo, L. Klotz, Biochem. Biophys. 595 (2016) 113–119. doi: 10.1016/j.abb.2015.06.024
H. Bozena, K. Marta, S. Sylvie, et al., Int. J. Mol. Sci. 18 (2017) 2209. doi: 10.3390/ijms18102209
H. Chuai, S. Zhang, H. Bai, et al., Eur. J. Med. Chem. 223 (2021) 113621. doi: 10.1016/j.ejmech.2021.113621
W. Zhou, P. Li, J. Liu, et al., Ind. Eng. Chem. Res. 59 (2020) 10763–10767. doi: 10.1021/acs.iecr.0c01147
X. Mao, P. Li, T. Li, et al., Chin. Chem. Lett. 31 (2020) 3276–3278. doi: 10.1016/j.cclet.2020.06.033
X. Xiao, Z. Shao, L. Yu, Chin. Chem. Lett. 32 (2021) 2933–2938. doi: 10.1016/j.cclet.2021.03.047
W. Ding, S. Wang, J. Gu, et al., Chin. Chem. Lett. 34 (2023) 108403. doi: 10.1016/j.cclet.2023.108403
B.K. Shimada, N. Alfulaij, L.A. Seale, Int. J. Mol. Sci. 22 (2021) 10713. doi: 10.3390/ijms221910713
M. Zhao, Q. Sun, M. Khogali, et al., Biol. Trace Elem. Res. 199 (2021) 4746–4752. doi: 10.1007/s12011-021-02603-7
M. Zhao, K. Wen, Y. Xue, et al., Animal 15 (2021) 100374. doi: 10.1016/j.animal.2021.100374
P. Surai, Br. Poult. Sci. 41 (2000) 235–243. doi: 10.1080/713654909
E. Delezie, M. Rovers, A. Van der Aa, et al., Poult. Sci. 93 (2014) 3083–3090. doi: 10.3382/ps.2014-04301
P. Surai, I. Kochish, Poult. Sci. 98 (2019) 4231–4239. doi: 10.3382/ps/pey406
V. Tufarelli, E. Cazzato, E. Ceci, et al., Biol. Trace Elem. Res. 173 (2016) 219–224. doi: 10.1007/s12011-016-0652-0
Y. Ohta, M. Kidd, T. Ishibashi, Poult. Sci. 80 (2001) 1430–1436. doi: 10.1093/ps/80.10.1430
N. Saeed, G. Jamshid, H. Shahin, et al., Int. J. Pept. Res. Ther. 24 (2018) 463–470. doi: 10.1007/s10989-017-9629-x
Z. Uni, P. Ferket, E. Tako, O. Kedar, Poult. Sci. 84 (2005) 764–770. doi: 10.1093/ps/84.5.764
X. Dong, Y. Jiang, M. Wang, et al., Poult. Sci. 92 (2013) 2118–2123. doi: 10.3382/ps.2013-03091
T. Pan, T. Liu, S. Tan, et al., Biol. Trace Elem. Res. 182 (2018) 364–372. doi: 10.1007/s12011-017-1110-3
X. Peng, Y. Cui, W. Cui, et al., Biol. Trace Elem. Res. 131 (2009) 33–42. doi: 10.1007/s12011-009-8345-6
S. Lee, H. Lillehoj, S. Jang, et al., Poult. Sci. 93 (2014) 1113–1121. doi: 10.3382/ps.2013-03770
L. Seale, H. Ha, A. Hashimoto, et al., Free Radic. Biol. Med. 127 (2018) 182–189. doi: 10.1016/j.freeradbiomed.2018.03.037
E. Zoidis, I. Seremelis, N. Kontopoulos, et al., Antioxidants 7 (2018) 66. doi: 10.3390/antiox7050066
J. Li, L. Zhang, Z. Yang, et al., Biol. Trace Elem. Res. 181 (2018) 340–346. doi: 10.1007/s12011-017-1049-4
M. El-Deep, K. Amber, S. Elgendy, et al., J. Anim. Physiol. Anim. Nutr. 104 (2020) 1392–1400. doi: 10.1111/jpn.13379
R. Krisnan, Y. Retnani, B. Tangendjaja, et al., Vet. World 14 (2021) 1109–1115.
L. Hou, H. Qiu, P. Sun, et al., Poult. Sci. 99 (2020) 6045–6054. doi: 10.1016/j.psj.2020.07.043
D. Yuan, X. Zhan, Y. Wang, Poult. Sci. 91 (2012) 936–942. doi: 10.3382/ps.2011-01921
X. Xiao, D. Yuan, Y. Wang, et al., Biol. Trace Elem. Res. 172 (2016) 201–208. doi: 10.1007/s12011-015-0541-y
I. Placha, J. Takacova, M. Ryzner, et al., Br. Poult. Sci. 55 (2014) 105–114. doi: 10.1080/00071668.2013.873772
Y. Ma, B. Cheng, Y. Li, et al., Biol. Trace Elem. Res. 200 (2021) 3785–3797.
G. Chen, J. Wu, C. Li, J. Anim. Physiol. Anim. Nutr. 98 (2014) 747–754. doi: 10.1111/jpn.12136
Y. Wang, B. Xu, Anim. Feed Sci. Technol. 144 (2007) 306–314.
M. Elahi, Y. Kong, B. Matata, Oxid. Med. Cell. Longev. 2 (2009) 259–269. doi: 10.4161/oxim.2.5.9441
D. Gessner, R. Ringseis, K. Eder, J. Anim. Physiol. Anim. Nutr. 101 (2017) 605–628. doi: 10.1111/jpn.12579
X. Wan, H. Ahmad, L. Zhang, et al., J. Sci. Food Agric. 98 (2018) 3715–3721. doi: 10.1002/jsfa.8879
W. Chen, L. Yue, W. Xia, Anal. Methods 10 (2018) 3534–3539. doi: 10.1039/C8AY00792F
G. Gaetani, A. Ferraris, M. Rolfo, et al., Blood 87 (1996) 1595–1599. doi: 10.1182/blood.V87.4.1595.bloodjournal8741595
J. Slot, H. Geuze, B. Freeman, et al., Lab. Investig. 55 (1986) 363–371.
I. Guido, A. Alessandro, F. Mariella, et al., Ital. J. Anim. Sci. 12 (2013) 131–133.

Figure 1 Flowchart of in ovo feeding of selenized glucose and incubated in neonatal broilers.

Figure 2 The effects of in ovo feeding of SeGlu on mRNA expression levels in the liver of chicks. a, b Different letters indicate significant differences among the three treatments (P < 0.05).
Table 1. Effects of in ovo feeding of selenized glucose (SeGlu) on hatching weight and selenium (Se) content on liver of chicks.
|
|
 下载: 导出CSV
下载: 导出CSV
Table 2. Effects of in ovo feeding of SeGlu on the GSH-system and thioredoxin-reductase enzyme in liver of chicks.
|
|
下载: 导出CSV
Table 3. Effects of in ovo feeding of SeGlu on free radical scavenging capacity in liver of chicks.
|
|
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们