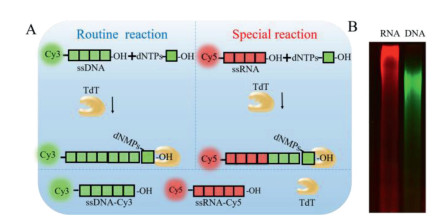
Figure 1.
The principle of TCED and TCER. (A) The illustration of ssDNA (in green) and ssRNA (in red) extension. (B) The image of 8% PAGE showed that 10−6 mol/L ssRNA and 10−6 mol/L ssDNA can be extended by TdT.
Single-stranded RNA as primers of terminal deoxynucleotidyl transferase for template-independent DNA polymerization
Houyu Han , Jingyu Cui , Dianming Zhou , Deping Hua , Weipan Peng , Mengyao Lin , Yating Zhang , Fangya Li , Xiaoqun Gong , Jianyu Zhang

Nucleic acid (DNA and RNA) plays a central role in the continuity of life. Due to the unique structural property and inherent ability [1, 2], DNA and RNA are usually widely used in drug delivery [3], biosensing [4], silencing [5, 6], and switching [7]. The key point for these applications lies in DNA or RNA extension synthesis [8-11]. To date, in addition to the synthesis of short-stranded nucleic acids by solid-phase synthesis, enzymatic synthesis was often used to synthesize high-molecular-weight nucleic acids, which catalyze the extension of 3′-hydroxy terminus (3′-OH) single-stranded DNA (ssDNA) and single-stranded RNA (ssRNA), and templates are usually essential for the extension of primers [12-16], which leads to the formation of DNA or chimeric RNA-DNA. For the most synthesis of chimeric RNA-DNA, templates were required. The template-independent synthesis of chimeric RNA-DNA was rarely reported. However, the requirement for using templates is not universal as exists a unique enzyme, denoted terminal deoxynucleotidyl transferase (TdT), which was first extracted from the calf thymus by F. J. Bollum [17-21]. TdT has been characterized as a template-independent polymerase using ssDNA as primers to generate random oligonucleotides [22-24]. Traditionally, the function of the TdT-catalyzed ssDNA extension has been preliminarily studied, including the influence of divalent metal cofactors for catalytic activity, and the crystal structures of the TdT [25-29].
This special template-independent property has led to numerous applications in biosensing [30, 31], gene engineering [10], and gene editing [11]. By examining the past publications, it is found that a few reports about the template-independent polymerases catalyzing the 3′-OH of RNA to add ribonucleotides, such as poly(U) polymerase and poly(A) polymerase [32, 33]. Yet, these enzymes were selected to use ribonucleotide substrates that greatly limit their capacity for diverse biotechnology applications. Polymerase theta (Pol θ) is essential for the double-strand break repair pathway alternative end-joining [34, 35]. Pomerantz group proved that the Pol θ can also perform one-step modification of ssRNA's 3′-termini in a manner of snap-back self-template-dependent extension activity [16, 36]. The limitation lies in the fact that Pol θ has a strong dependence on the structure of primers, requiring the 3′-end primer to form a pair with itself [16, 36].
In previous work, single-chain poly(T) was synthesized by TdT based on this trait to activate the self-assembly of copper nanoparticles to detect Parkinson's disease-related microRNA153–3p [37]. However, when all DNA can be effectively blocked, a false positive signal was still observed with a relatively high concentration of RNA in the process of the experiment. It is speculated that in addition to ssDNA, TdT could catalyze the extension of ssRNA by incorporating deoxyribonucleotide (dNTPs) to form chimeric RNA-DNA. Therefore, according to relevant literature, it is known that TdT can be extended with ssRNA as primers, but there are only several related kinds of literature [38, 39]. In this work, TdT polymerization reactions with ssRNA and ssDNA as primers were contrasted in detail to synthesize chimeric RNA-DNA, which has a wide range of applications such as gene targeting, PCR primers, and biosensors [40-42]. If chimeric RNA-DNA can be constructed by TdT, it will open a new window in which ssRNA extends DNA by adding dNTPs in the absence of templates.
Therefore, we detailed studied the extension performance with ssRNA as primers, by comparison with ssDNA, the TdT for its ability to perform DNA synthesis without templates using ssDNA and ssRNA primers was uncovered. In this work, considering that chimeric RNA-DNA has demonstrated extensive growth and shown increasing importance in medicine, we were devoted to exploring the function of TdT-catalyzed 3′-OH ssRNA extension (TCER), targeting to find out the detailed performance of both TCER and TdT-catalyzed 3′-OH ssDNA extension (TCED).
To our best knowledge, simple and efficient methods to detect TdT-catalyzed profiles are lacking, and the gel electrophoresis technique is the most commonly used [16, 43, 44]. To visually distinguish the extension effect of TdT on different primers, we finally chose fluorophores as labels for characterization. Herein, as shown in Fig. 1A, to effectively distinguish between ssRNA and ssDNA, the cyanine dyes Cy3 and Cy5 were used to label corresponding ssDNA and ssRNA with different colors in polyacrylamide gel electrophoresis (PAGE): the 5′-PO4-ssDNA was labeled with Cy3 (ssDNA-Cy3) showing green fluorescence and the 5′-PO4-ssRNA was colored with Cy5 (ssRNA-Cy5) with red fluorescence [45-47]. To verify the feasibility of TCER and TCED, the principles were illustrated in Fig. 1A, and the image of 8% PAGE showed that ssRNA and ssDNA can be extended by TdT (Fig. 1B). Control experiments have shown that TdT would catalyze the extension of DNA and RNA only when all components, including dNTPs and TdT, are present (Fig. S1 in Supporting information). The sequences of ssDNA and ssRNA were listed in Table S1 (Supporting information). It is suggested that TdT can not only catalyze ssDNA-primed DNA polymerization but also can catalyze ssRNA-primed DNA polymerization. In order to further illustrate the extension performance of TdT, several ssRNA with different sequences were randomly selected to compare their extension efficiency (Table S1). As microRNA 122 was widely used as a disease marker with high extension efficiency [48, 49], it was chosen to study the performance of TdT extension RNA (Fig. S2 in Supporting information). In addition, normal primers rather than specified primers which 3′ end can be paired with itself were designed (Fig. S3 in Supporting information) to test the possibility of RNA extension [16].
When different concentrations of ssRNA and ssDNA were extended by TdT (Figs. 2A–C), it is clear that with a higher concentration of primers, fluorescence intensity became stronger with a wider size distribution of polymerization products. All unextended primers have migrated out of the electrophoretic lane, and the counted fluorescence intensity represents the fluorescence intensity of the extended primers, i.e. the whole lane. A near-perfect linear trend was observed for the plot of fluorescence intensity with the concentration of nucleic acid (Fig. 2B). The slope for RNA primers is lower than for DNA primers, indicating that the catalytic efficiency of TCER was poorer than TCED. The fluorescence intensity was correlated with the concentrations of ssDNA and ssRNA, enabling us to quantitatively analyze Cy5 and Cy3 fluorescence intensity, and standard curves of different concentrations of ssRNA and ssDNA with fluorescence intensity were fitted (Fig. S4 in Supporting Information). Through calculation (detail method is shown in Supporting information and data are shown in Figs. S5 and S6 in Supporting Information), it is concluded that when the concentration of ssDNA or ssRNA increased, the percentage of extended primers in the reaction decreased: the case for ssRNA decreased sharply from around 47% to 12%; while in the case of ssDNA, the deduction is limited with more than about 85% of extended primers in the reaction (Fig. 2C). Furthermore, when the amount of enzyme in the reaction system was increased to 10 folds, the proportion of extended ssRNA in the reaction was not significantly increased, suggesting that ssDNA is more easily captured by TdT (Fig. S7 in Supporting information). From Fig. 2A and Fig. S6, it is interesting to find out that the extension products of ssRNA were concentrated at the upper part of the lane no matter with a high or low concentration of ssRNA primers, indicating that once ssRNA was captured by TdT and extended to form chimeric RNA-DNA, which may continue to be extended easily by TdT. This was more probable to be captured by TdT again than ssRNA itself, which may be due to its cellular function as a DNA polymerase [50]. If we looked more detailly at the molecular weight (MW) of the reaction products, it is found that the products of TCER stick to the high MW part (> 1200 bp), and gradually extend to the lower MW part as more ssRNA primers are available (Fig. S6C). In the case of ssDNA primers, as the concentration of primers increased, the products were more evenly distributed on the lane (Fig. 2A and Fig. S6D). When the concentration of primers is low, the polymerization product prefers a large MW (> 1200 bp) product. With ssDNA higher than 5 × 10−7 mol/L, products were concentrated on the "100–500 bp" or "500–1200 bp" parts, consistent with previous reports [43]. Therefore, TCER could effectively solve the problem that current methods are difficult to produce long-chain nucleic acids and generate products with high molecular weight, which can be used in synthetic biology or to produce DNA tails to protect RNA from deterioration [51].
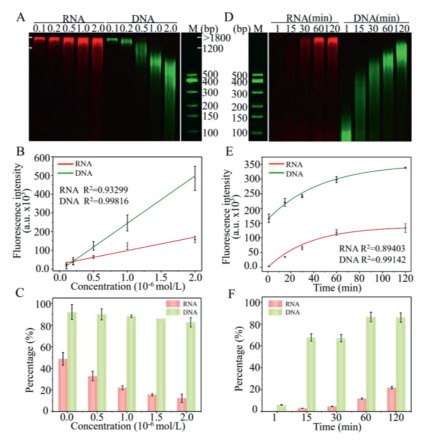
By comparing the extension efficiency of different concentrations of ssDNA and ssRNA, we continued to explore the effect of reaction time on TdT catalysis (Figs. 2D-F). As the reaction time increased, ssRNA and ssDNA primers continued to be extended with obviously different extension characteristics. The products of TCER were still concentrated in the upper part of the lane no matter how long the reaction time was. With TdT reflected in the relatively uniform extension using ssDNA as primers, the total MW gradually increased with extended reaction time (Fig. 2D). Either TCER or TCED, the fluorescence intensity significantly enhanced as the reaction time increased (Fig. 2E). At 120 min time point, about 86% of ssDNA primers were extended, indicating that ssDNA had a relatively equal chance of being captured and extended by TdT. At the same time, only around 22% of ssRNA primers were extended (Fig. 2F and Fig. S8 in Supporting information). Through quantitative analysis of fluorescence intensity and MW at different positions on the gel band, we were able to estimate the number of base pairs participating in the reaction at different time points. With these parameters, the reaction rates for TCER and TCED were estimated to be (341 ± 12) × 10−9 mol min−1 mg−1 and (983 ± 37) × 10−9 mol min−1 mg−1, respectively (Fig. S8). This result indicated that the catalytic efficiency of TCER was about 3 times lower than TCED. The reason was that around 22% of ssRNA primers participated in the reaction much less than 86% of ssDNA in 120 min, but the growth trend of ssRNA primers was toward larger MW products (mostly > 1200 bp). The MW of single-stranded DNA-DNA was much smaller than chimeric RNA-DNA, leading to the final catalytic efficiency of TCER not much less than TCED. In general, it is proved that TdT also had high catalytic activity for ssRNA primers, which can be used for the template-independent synthesis of chimeric RNA-DNA and is widely adopted in biomedical research and applications.
To better understand the extension preference through different substrates, we proceeded to investigate the effects of different substrates (Figs. 3A and B), and the two gels and DNA marker gel run at the same time in the same gel box (Fig S9 in Supporting information). These results demonstrated that both TCER and TCED show a clear preference for different substrates, and the proportion of extended primers in the reaction is different with different substrates (Figs. 3C and Fig. S10 in Supporting information). dATP and dTTP were regarded as poor substrates for TCER. Substrates dCTP, dGTP and dNTPs showed relatively good activity for ssRNA primers, mainly distributed in the "100–500 bp" part. And there are two reasons for ssDNA extension with dATP affording a quite narrow band. On the one hand, it may be the interactions of the negatively-charged triphosphate moieties with positively-charged amino acids in the polymerase's active site [25]. On the other hand, it may be related to divalent metal cations (Co2+) that were used in the experiment to promote the binding of pyrimidine bases [25]. By calculating the concentration of extended ssRNA and ssDNA in the reaction according to fluorescence intensity based on the standard curve (Figs. S4, S5 and S10), the number of base pairs in the products could be obtained based on the DNA marker. The results showed that the preferred order of substrates for extension efficiency of TCER was dNTPs > dCTP > dGTP > dATP > dTTP and the order of TCED for preferred substrate was dNTPs > dTTP > dATP > dCTP > dGTP (Fig. 3D and Fig. S10), which may be related to the conformational change of the active center of TdT upon binding to the base and the divalent metal cation [25]. it is found that the MW distribution was also substrate-dependent: dATP prefers to form the "500–1200 bp" product, and dTTP, dGTP, and dCTP as substrates mainly grow in "100–500 bp" (Figs. S10F and G).
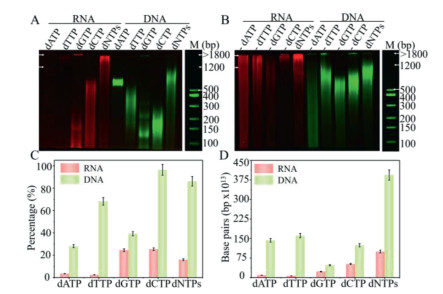
In addition to single substrate performance, we further carried out single substrate knock-out experiments to test the polymerization characteristics of TCER and TCED (Fig. 3B and Fig. S11A in Supporting information). With the absence of dATP (-dATP), the products of TCED were evenly distributed in the lane, indicating that the dATP as the substrate in TCED is critical in determining the high MW characteristics of the products. When dNTPs lacked dCTP or dGTP (-dCTP or -dGTP), TCER showed poor growth with the products mainly concentrated on "500–1200 bp" and "1200–1800 bp" parts (Figs. S11B and C in Supporting information). The results suggested that dCTP and dGTP played a major role in the extension. In the process of TCED, the lack of any kind of deoxynucleotide alone had little effect on the extension, and the products were almost distributed in "100–500 bp" and "500–1200 bp" parts (Figs. S11D and E in Supporting information). The choice of substrates also had a significant effect on the behavior of polymerization, overall, data in Figs. 3A and B demonstrated that TdT exhibited a strong preference for different substrates providing a valid reference for the growth of nucleic acid strands of different deoxynucleotides in the absence of templates.
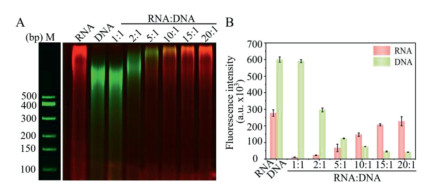
Since TdT could catalyze the polymerization using both ssDNA and ssRNA primers, it would be necessary to understand the competitive behavior between ssDNA and ssRNA. Here, different ssRNA/ssDNA primer ratios ranging from 1:1 to 20:1 were tested with a constant concentration of ssRNA (10−6 mol/L) (Fig. 4 and Fig S12 in Supporting information). From Figs. 4A and B, it is clear that when the ratio is less than 5, TdT prefers to extend ssDNA. As the ssDNA concentration decreased with ssRNA/ssDNA ratio higher than 10, ssRNA was preferentially captured and extended by TdT. Although the exact affinity of nucleic acid to TdT is unknown, it is estimated that the ssDNA could bind tighter than ssRNA, because TdT is a DNA polymerase. TdT can catalyze the formation of polynucleotides at 3′-OH of ssRNA with higher concentration which can be preferentially captured and extended by TdT. This also explained the background of our previous work [37].
TdT is well known for its ability to add dNTPs on 3′-OH of ssDNA with the characteristic of template-independent. In this process of studying TdT, we described that TdT has strong catalytic activity for ssRNA as primers without any templates in detail. It is proposed that ssRNA can be extended by TdT to produce DNA tails to protect RNA from deterioration. When ssRNA is extended by TdT to form chimeric RNA-DNA, high MW products at 3′-OH of ssRNA were preferred. The catalytic efficiency of TCED was about 3 times higher than TCER. TCER was found to depend on substrates with the order of dNTPs > dCTP > dGTP > dATP > dTTP. The presented study offers a promising launching point for the development of a practical template-independent extension of 3′-OH of ssRNA technology. Finally, it is believed that understanding the performance of TCER and TCED will help to design the strategy to synthesize chimeric DNA on 3′-OH of ssRNA. The crystal structure of TCED had been understood [25], while we are now studying the crystal structure of TCER to detail the interaction of TdT with ssRNA, trying to fully understand the extending mechanism shortly.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported by the National Natural Science Foundation of China (NSFC, Nos. 21927814 and 21772143 to J.Y. Zhang), and the National Science Foundation of Tianjin (Nos. 20YDTPJC00090, 19ZXDBSY00070 and 20YFZCSY00990 to X.Q. Gong).
Supplementary material associated with this article can be found, in the online version, at doi:
J.D. Watson, F.H.C. Crick, Nature 171 (1953) 737–738. doi: 10.1038/171737a0
V.V. Demidov, Drug Discov. Today 6 (2001) 178–179. doi: 10.1016/S1359-6446(00)01635-4
M. Kwak, A. Herrmann, Chem. Soc. Rev. 40 (2011) 5745–5755. doi: 10.1039/c1cs15138j
R. Gasparac, B.J. Taft, M.A. Lapierre-Devlin, et al., J. Am. Chem. Soc. 126 (2004) 12270–12271. doi: 10.1021/ja0458221
A.J. Hamilton, Science 286 (1999) 950–952. doi: 10.1126/science.286.5441.950
S.M. Elbashir, J. Harborth, W. Lendeckel, et al., Nature 411 (2001) 494–498. doi: 10.1038/35078107
B.J. Tucker, R.R. Breaker, Curr. Opin. Struct. Biol. 15 (2005) 342–348. doi: 10.1016/j.sbi.2005.05.003
M. Chen, X. Gong, J. Chang, et al., Anal. Chem. 92 (2020) 13336–13342. doi: 10.1021/acs.analchem.0c02642
M. Liu, J. Song, C. Dong, et al., ACS Nano 8 (2014) 5564–5573. doi: 10.1021/nn5007418
B. Gao, Q. Sun, Nat. Commun. 12 (2021) 2689. doi: 10.1038/s41467-021-22894-7
R.A. Gier, Z. Cao, J. Shi, et al., Nat. Commun. 11 (2020) 3455. doi: 10.1038/s41467-020-17209-1
M. Safak, F.E. Alemdaroglu, Y. Li, et al., Adv. Mater. 19 (2007) 1499–1505. doi: 10.1002/adma.200700240
G.J. Hafner, M.R. Stafford, P.M. Giffard, et al., BioTechniques 30 (2001) 852–867. doi: 10.2144/01304rr03
E.A. Manrao, I.M. Derrington, A.H. Laszlo, et al., Nat. Biotechnol. 30 (2012) 349–353. doi: 10.1038/nbt.2171
M. Brownstein, BioTech. 20 (1996) 1004–1006. doi: 10.2144/96206st01
C. Thomas, T. Rusanov, T. Hoang, et al., Nucl. Acids Res. 47 (2019) 3272–3283. doi: 10.1093/nar/gkz029
F.J. Bollum, Blood 54 (1979) 1203–1215. doi: 10.1182/blood.V54.6.1203.1203
F.J. Bollum, J. Biol. Chem. 234 (1959) 2733–2734. doi: 10.1016/S0021-9258(18)69770-4
F.J. Bollum, V.R. Potter, J. Biol. Chem. 233 (1958) 478–482. doi: 10.1016/S0021-9258(18)64787-8
E.A. Motea, A.J. Berdis, Biochim. Biophys. Acta 1804 (2010) 1151–1166. doi: 10.1016/j.bbapap.2009.06.030
K. Ramadan, G. Maga, U. Hübscher, et al., J. Biol. Chem. 339 (2004) 395–404.
J.B. Boulé, C.J. Papanicolaou, C. Papanicolaou, Biol. Chem. 276 (2001) 31388–31393. doi: 10.1074/jbc.M105272200
R. Roychoudhury, E. Jay, R. Wu, Nucl. Acids Res. 3 (1976) 863–877. doi: 10.1093/nar/3.4.863
I. Sarac, M. Hollenstein, ChemBioChem 20 (2019) 860–871. doi: 10.1002/cbic.201800658
M. Delarue, Embo J. 21 (2014) 427–439.
L. Chang, F.J. Bollum, J. Biol. Chem. 247 (1967) 536–542.
L. Chang, F.J. Bollum, J. Biol. Chem. 265 (1990) 17436–17440. doi: 10.1016/S0021-9258(18)38181-X
J. Gouge, P. Beguin, M. Delarue, et al., J. Mol. Biol. 425 (2013) 4334–4352. doi: 10.1016/j.jmb.2013.07.009
Y. Zhang, J.H. Liang, X.W. Tao, et al., Biotechnol. Bull. 35 (2019) 161–169.
Z. Liu, X. Luo, L. Zhu, et al., Anal. Chem. 89 (2017) 1892–1899. doi: 10.1021/acs.analchem.6b04360
L. Wang, Y. Pan, G. Li, et al., ACS Appl. Mater. Interfaces 12 (2020) 322–329. doi: 10.1021/acsami.9b18869
G. Cao, J.N. Sarkar, Proc. Natl. Acad. Sci. U. S. A. 89 (1992) 10380–10384. doi: 10.1073/pnas.89.21.10380
Y. Zhou, H. Yin, X. Zhang, et al., Biosens. Bioelectron. 79 (2016) 79–85. doi: 10.1016/j.bios.2015.12.009
T. Kent, A.Y. Ozdemir, et al., Nat. Struct. Mol. Biol. 22 (2015) 230–237. doi: 10.1038/nsmb.2961
S.H. Chan, A.M. Yu, R.S. Hawley, PLoS Genet. 6 (2010) 1–16.
T. Kent, P.A. Gomez, A. Sfeir, R.T. Pomerantz, eLife 5 (2016) 1–25.
J. Cui, H. Han, J. Chang, et al., ACS Appl. Mater. Interfaces 12 (2020) 34130–34136. doi: 10.1021/acsami.0c06032
B. Zhao, Z. Gong, Y. Jin, et al., Acta Biochim. Biophys. Sin. 44 (2012) 129–135. doi: 10.1093/abbs/gmr115
I.D. Vilfan, W. Kamping, M. van den Hout, et al., Nucl. Acids Res. 19 (2007) 6625–6639.
A.F. El-Yazbi, G.R. Loppnow, Anal. Chem. 85 (2013) 4321–4327. doi: 10.1021/ac301669y
H. Miyachi, S. Hoshina, I. Karube, et al., Anal. Chim. Acta 407 (2000) 1–10. doi: 10.1016/S0003-2670(99)00779-5
K. Hanseop, L. Wi-Jae, O. Yeounsun, et al., Nucl. Acids Res. 48 (2020) 8601–8616. doi: 10.1093/nar/gkaa605
L. Tang, L.A. Navarro, A. Chiloti, S. Zauscher, Angew. Chem. Int. Ed. 56 (2017) 6778–6782. doi: 10.1002/anie.201700991
V. Rosemeyer, A. Laubrock, R. Seibl, Anal. Biochem. 224 (1995) 446–449. doi: 10.1006/abio.1995.1068
W. Lee, P. Hippel, A.H. Marcus, Nucl. Acids Res. 42 (2014) 5967–5977. doi: 10.1093/nar/gku199
Y. Fu, J.R. Lakowicz, Anal. Chem. 78 (2006) 6238–6245. doi: 10.1021/ac060586t
A. Iqbal, S. Arslan, B. Okumus, et al., Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 11176–11181. doi: 10.1073/pnas.0801707105
X. Zhu, X. Wang, S. Li, et al., ACS Sen. 6 (2021) 881–888. doi: 10.1021/acssensors.0c01984
H. Šípová, S. Zhang, A.M. Dudley, et al., Anal. Chem. 82 (2010) 10110–10115. doi: 10.1021/ac102131s
J. Loc'H, M. Delarue, Curr. Opin. Struct. Biol. 53 (2018) 22–31. doi: 10.1016/j.sbi.2018.03.019
Y.J. Chen, B. Groves, R.A. Muscat, G. Seelig, Nat. Nanotechnol. 10 (2015) 748–760. doi: 10.1038/nnano.2015.195

Figure 1 The principle of TCED and TCER. (A) The illustration of ssDNA (in green) and ssRNA (in red) extension. (B) The image of 8% PAGE showed that 10−6 mol/L ssRNA and 10−6 mol/L ssDNA can be extended by TdT.

Figure 2 The TCER and TCED performance at different primers concentrations and different reaction times. Two-channel (Cy5 and Cy3) analysis of the TCER and TCED performance at different primers concentrations (A) and different reaction times (D), respectively by PAGE images (ssRNA and ssDNA reaction concentrations increase from 10−7 mol/L to 2 × 10−6 mol/L and the reaction time increases from 1 min to 120 min). Relationship between the fluorescence intensity and the concentration of ssRNA and ssDNA primers (B) and the reaction time (E). The proportion of extended ssRNA and ssDNA primers in the reaction in different primers concentrations (C) and different reaction times (F). n = 3; error bars represent standard deviation.

Figure 3 The performance of TCER and TCED with different substrates. (A, B) Two-channel (Cy5 and Cy3) analysis of the TCER and TCED performance with different substrates, respectively by PAGE images. (A) TCER and TCED are from lanes 1 to 5 and from lanes 6 to 10 using dATP, dTTP, dCTP, dGTP, and dNTPs as substrates. (B) TCER and TCED are from lane 1 to 5 and from lanes 6 to 10 using -dATP, -dTTP, -dCTP, -dGTP and dNTPs as substrates. (C) The proportion of extended ssDNA and ssRNA primers in the reaction with different substrates (A). (D) The number of base pairs of TCER and TCED products with different substrates (A). n = 3; error bars represent standard deviation.

Figure 4 The characteristics of ssRNA and ssDNA coexisting in the same reaction system catalyzed by TdT. (A) Two-channel (Cy5 and Cy3) analysis of ssRNA and ssDNA coexisting in the same reaction system catalyzed by TdT by PAGE images (Lane 1: 10−6 mol/L RNA, Lane 2: 10−6 mol/L DNA, Lane 3: 10−6 mol/L RNA + 10−6 mol/L DNA, Lane 4: 10−6 mol/L RNA + 5 × 10−7 mol/L DNA, Lane 5: 10−6 mol/L RNA + 2 × 10−7 mol/L RNA, Lane6: 10−6 mol/L + 10−7 mol/L DNA, Lane 7: 10−6 mol/L + 6 × 10−8 mol/L DNA, Lane 8: 10−6 mol/L + 5 × 10−8 mol/L DNA). (B) Fluorescence intensity of ssRNA and ssDNA coexisting in the same reaction system catalyzed by TdT. n = 3; error bars represent standard deviation.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载: