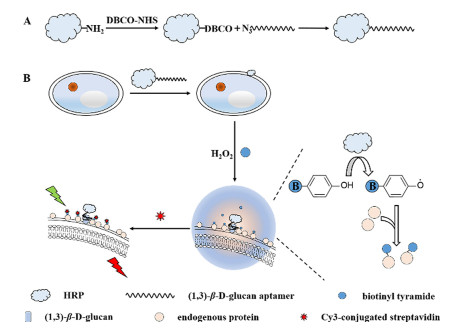
Figure 1.
(A) Procedure of preparation of aptamer-conjugated HRP. (B) Schematic illustration of specific recognition of aptamer to (1, 3)-β-D-glucan and tyramide signal amplification technology for the identification of fungi.
In situ fluorescence imaging of fungi via (1, 3)-β-D-glucan aptamer and tyramide signal amplification technology
Pengfei Zuo , Feng Gong , Yixia Yang , Xinghu Ji , Zhike He

Fungal infections have aroused great concern due to their increasing infection incidences and various types of pathogenic fungi that threaten the human health [1]. Fungi are a class of microorganisms that consist of a tough cell wall, a nucleus and complete organelles. The cell wall of fungi is the main difference compared with other cells, which comprises chitin, β-glucan, mannan, glycoprotein and other polysaccharides and proteins. They act on maintaining the normal growth and physiological functions of fungal cells [2]. Thus, these biomolecules are potentially useful in identifying fungi.
As the diagnostic gold standards for many fungal infections, culture-based methods possess high accuracy but low sensitivity and time-consuming procedures [3]. By contrast, fluorescence imaging is a more rapid and simple technology than culture-based methods. It could be applied in detecting biomolecules in living cells [4, 5]. Typically, optical brightener non-specifically binds to polysaccharides of cell wall of fungi such as chitin and glucan. It could emit strong blue fluorescence with the excitation of ultraviolet light that proves the existence of fungi [6]. However, cellulose and other polysaccharides adhered to the human skin accidentally disturb the diagnostic results. To handle this, fluorescent probes specifically conjugating to the biomarkers of cell wall of fungi instead of optical brightener are taken into account, which improve the accuracy and specificity of fungi detection [7]. Recently, several specific fluorescent probes have been developed for identifying fungi. Shao applied fluorescein-labeled chitinase binding to chitin to diagnose fungal infections [8]. Baibek employed fluorescein conjugated antifungal polyene amphotericin B by binding to ergosterol, a component that is specific to the fungal cell membrane, to identify microbial species in a co-culture of fungi and bacteria [9].
Glucans are water-insoluble structural cell wall components of most fungi, plants and some bacteria. In the cell wall of fungi, the primary constituent of glucans is (1, 3)-β-D-glucan, which presents the diagnostic criteria of invasive fungal infections. (1, 3)-β-D-glucan is a specific biomarker for fungi [10, 11]. In the meanwhile, aptamers are the artificial single-stranded DNA or RNA sequences that fold into secondary and tertiary structures. They could bind to certain targets with extremely high specificity such as small molecules, proteins even single cells [12, 13]. Compared with antibodies, enzymes or other binding proteins, aptamers are more stable, easily available and modifiable. Aptamers have been widely applied in cell imaging, point-of-care diagnosis, precision medicine and even tumor therapy [14-17]. And Low developed the (1, 3)-β-D-glucan aptamer for the detection of fungi [18]. Hence, the aptamer specifically binding to (1, 3)-β-D-glucan is potentially efficient for identifying fungi.
For highly sensitive sensing of fungi, a signal amplification technology is introduced to combine with aptamers. Tyramide signal amplification technology (TSA) is a horseradish peroxidase (HRP) mediated signal amplification method. It utilizes HRP to convert a tyramide substrate into a reactive radical that covalently tags neighboring proteins with the presence of hydrogen peroxide (H2O2) [19, 20]. HRP is usually targeted to specific biomarkers via conjugation to a protein or antibody for a signal amplification [21]. TSA technology has been widely used in immunohistochemistry, fluorescence in situ hybridization, ELISA, and proteomic profiling of cells for a higher sensitivity and accuracy [22-26].
In this work, taking advantage of aptamers specifically recognizing (1, 3)-β-D-glucan and TSA technology, we developed a specific and sensitive method for the in situ fluorescence imaging of fungi (Fig. 1). The yeast from baking ingredients was selected as the model of fungi. As shown in Fig. 1A, HRP was firstly modified with dibenzocyclooctyne (DBCO). HRP-DBCO was then conjugated with aptamer modified with azide at the 5′ end quickly and gently via click chemistry [27]. One end of aptamer modified with HRP was successfully assembled. And then aptamer-conjugated HRP bound to the cell wall of yeast through the specific recognition of aptamer to (1, 3)-β-D-glucan. With the addition of biotinyl tyramide and hydrogen peroxide, HRP could catalyze biotinyl tyramide into biotinyl radical that tags neighboring proteins with biotin in a few minutes [28]. Multiple Cy3-conjugated streptavidin were capable of binding to yeast with a fluorescence signal simplification, which was depicted in Fig. 1B. By the red fluorescence signal of multiple Cy3, the existence and characteristic shape of fungi could be observed by the fluorescence microscope with the excitation of 546 nm. Based on the high specificity of aptamer and signal amplification of TSA technology, a specific and sensitive in situ fluorescence imaging method was constructed to detect fungi with the morphology information.
To verify the successful ligation of HRP with aptamer via click reaction, UV–vis absorption spectra and zeta potentials were investigated. As shown in Fig. S1 (Supporting information), the UV absorption spectrum of aptamer-conjugated HRP contained two peaks at 259 nm and 404 nm, which represent the characteristic UV absorption peak of aptamer at 258 nm and HRP at 403 nm. This indicated the conjugation of aptamer and HRP. As Shown in Fig. S2 (Supporting information), the zeta potential of HRP (−4.79 eV) increased to −3.71 eV after the modification of DBCO. The zeta potential of aptamer-conjugated HRP (−21.8 eV) was larger than that of N3-aptamer (−29.4 eV) and smaller than that of HRP-DBCO (−3.71 eV). These results also confirmed the successful ligation of aptamer and HRP through click chemistry.
Meanwhile, yeast was incubated with aptamer-conjugated HRP and centrifuged several times to remove the uncombined aptamer-conjugated HRP. 3, 3′, 5, 5′-Tetramethylbenzidine (TMB) liquid substrate was then respectively added into yeast-HRP, supernatant of centrifugation and only yeast to demonstrate the combination of aptamer-conjugated HRP with yeast. As shown in Fig. S3 (Supporting information), it was obvious that the color of supernatant gradually changed from deep yellow to transparent (washing times: 1–4 times), implying that free aptamer-conjugated HRP was removed. The color of yeast-HRP turned from colorless to navy blue proving the catalytic ability and existence of HRP, while the yeast without incubation with aptamer-conjugated HRP remained colorless. Above results revealed that aptamer-conjugated HRP successfully bound to the yeast.
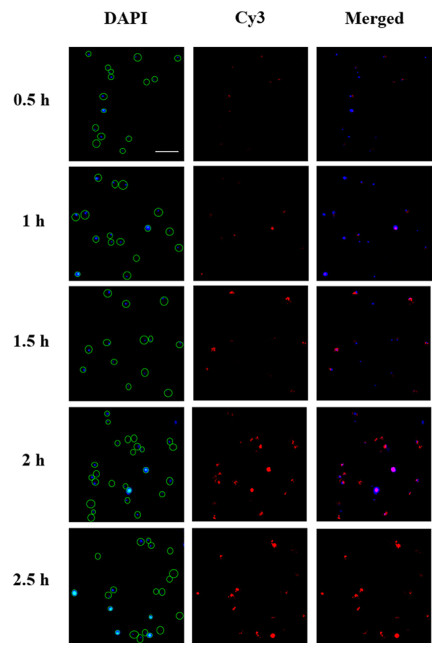
In order to visualize the fungi accurately, the incubation time of yeast with aptamer was taken into account. The yeast was conjugated with biotin-aptamer for different times (0.5 h, 1 h, 1.5 h, 2 h and 2.5 h). As shown in Fig. 2, nucleus labeled with 4′, 6-diamidino-2-phenylindole (DAPI) functionalizing as location of yeast emitted blue fluorescence with the excitation of 365 nm. With the increase of labeling time (0.5–2 h), most fungi were labeled with Cy3 and emitted red fluorescence. Finally, 2 h was chosen as the optimized incubation time.
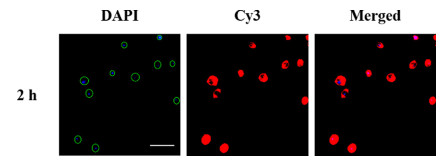
Herein, TSA technology was introduced to increase the accuracy and sensitivity of fluorescence imaging. With aptamer-conjugated HRP recognizing (1, 3)-β-D-glucan of yeast, biotinyl tyramide could be catalyzed into biotinyl radical by HRP that tags neighboring proteins on the cell wall with biotin in the presence of hydrogen peroxide. Then, multiple Cy3-conjugated streptavidin bound to yeast through high affinity for biotin with fluorescence signal amplification. As shown in Fig. 3, with the location of yeast by DAPI, more bright and intense red fluorescence of Cy3 could be observed. The fluorescence microscope images of 2 h described the shape of yeast preferably and confirmed the success of fluorescence signal amplification with less background interference. As a comparison, aptamer conjugated with Cy3 in Fig. 2 without signal amplification could not fully provide the morphology and diagnostic information for identifying fungi. Moreover, even incubated for only 0.5 h (Fig. S4 in Supporting information), the shape of yeast was clearly imaged, while in Fig. 2 almost no red fluorescence could be observed of 0.5 h. With TSA technology, the incubation time could be greatly reduced for an effective imaging.
To gain further insight into the specificity of aptamer to (1, 3)-β-D-glucan, a random DNA sequence was used to replace (1, 3)-β-D-glucan aptamer and the result was depicted in Fig. S5 (Supporting information). Different from (1, 3)-β-D-glucan aptamer, almost no red fluorescence could be observed with the addition of Cy3-conjugated streptavidin in the field of fluorescence microscope, indicating the specific interaction of aptamer with (1, 3)-β-D-glucan. Meanwhile, Escherichia coli, a typical symbol of bacteria, was selected as the control group. As shown in Fig. S6 (Supporting information), E. coli was incubated with DAPI for location. With the addition of Cy3-conjugated streptavidin, alomost no red fluorescence could be observed. This demonstrated that bacteria was not be recognized by (1, 3)-β-D-glucan aptamer. Fungi could be detected with (1, 3)-β-D-glucan aptamer specifically even in the presence of bacteria. Fluorescence spectrum was also carried out to prove the specificity. As shown in Fig. S7 (Supporting information), the fluorescence of 6-carboxyfluorescein-aptamer (FAM-aptamer) was quenched in the presence of graphene oxide (GO), as GO could adsorb dye-labeled aptamer via hydrophobic and π-π stacking interactions [29, 30]. The fluorescence of FAM-aptamer was recovered with the addition of β-D-glucan, while in the presence of mannan, laminarin, glucose or d-trehalose dihydrate, the fluorescence of FAM-aptamer was not recovered. This further demonstrated the specificity of aptamer to (1, 3)-β-D-glucan.
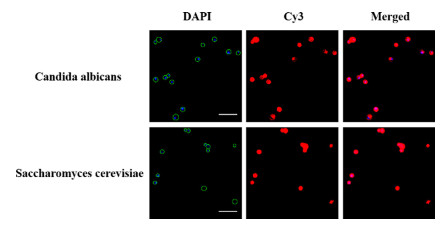
To evaluate the reliability and applicability of this proposed fluorescence signal amplification method for the detection of fungi, cultured candida albicans and saccharomyces cerevisiae were tested. As depicted in Fig. 4, the ellipsoidal shapes of candida albicans and saccharomyces cerevisiae could be significantly observed by fluorescence microscope, indicating a great potential of the proposed method for identifying fungi. The morphology and diagnostic information of fungi were distinctly expressed by TSA technology. Compared with other reported methods for fungi detection, the combination of (1, 3)-β-D-glucan aptamer and TSA technology possesses high specificity and sensitivity. (Table S2 in Supporting information). Thus, this method proved to be an effectively universal method for the detection of fungi.
In conclusion, this work demonstrated a specific and sensitive in situ fluorescence imaging method for the detection of fungi. The accurate and specific detection was attributed to the recognition of aptamer to (1, 3)-β-D-glucan. And with the advantageous of TSA technology, fluorescence signal could be amplified greatly which accelerates the procedure of observation and detection of fungi. By the application of this method, the combination of aptamer and TSA technology is a promising tool for the detection of fungi, bacteria and even eukaryotic cell with the virtue of biomarkers.
There are no conflicts to declare.
This work was supported by the National Natural Science Foundation of China (Nos. 22174102, 21974101, 22174101) and the Sino-German rapid response funding call for Covid-19 related research (No. C-0008).
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2022.02.064.
A.F. Chapuis, E.R. Ballou, D.M. MacCallum, J. Fungi 5 (2019) 29. doi: 10.3390/jof5020029
I. Geoghegan, G. Steinberg, S. Gurr, Trends Microbiol. 25 (2017) 957–967. doi: 10.1016/j.tim.2017.05.015
D. Limmathurotsakul, K. Jamsen, A. Arayawichanont, et al., PLoS One 5 (2010) e12485. doi: 10.1371/journal.pone.0012485
T. Kowada, H. Maeda, K. Kikuchi, Chem. Soc. Rev. 44 (2015) 4953–4972. doi: 10.1039/C5CS00030K
S.S. Long, Q.L. Qiao, L. Miao, Z.C. Xu, Chin. Chem. Lett. 30 (2019) 573–576. doi: 10.1016/j.cclet.2018.11.031
R. Ruchel, M. Schaffrinski, K.R. Seshan, G.T. Cole, Med. Mycol. 38 (2000) 231–237. doi: 10.1080/mmy.38.3.231.237
C. Zhao, L. Mendive-Tapia, M. Vendrell, Arch. Biochem. Biophys. 661 (2019) 187–195. doi: 10.1016/j.abb.2018.11.018
J. Shao, Y.G. Song, Y.B. Zhou, et al., Med. Mycol. 58 (2020) 66–70. doi: 10.1093/mmy/myz035
A. Baibek, M. Ucuncu, B. Short, et al., Chem. Commun. 57 (2021) 1899–1902. doi: 10.1039/d0cc08177a
C.M. de Sousa Lacerda, I.M. Ferreira, S.R. dos Santos, et al., Nucl. Med. Biol. 46 (2017) 19–24. doi: 10.1016/j.nucmedbio.2016.11.008
D.L. Williams, Mediators Inflamm. 6 (1997) 247–250. doi: 10.1080/09629359791550
A.D. Ellington, J.W. Szostak, Nature 355 (1992) 850–852. doi: 10.1038/355850a0
A.B. Iliuk, L.H. Hu, W.A. Tao, Anal. Chem. 83 (2011) 4440–4452. doi: 10.1021/ac201057w
Y.Y. Fan, L. Li, M. Lu, H.B. Si, B. Tang, Chem. Commun. 55 (2019) 4043–4046. doi: 10.1039/c9cc00244h
T. Wang, L.M. Chen, A. Chikkanna, et al., Theranostics 11 (2021) 5174–5196. doi: 10.7150/thno.56471
L.L. Wu, Y.D. Wang, X. Xu, et al., Chem. Rev. 121 (2021) 12035–12105. doi: 10.1021/acs.chemrev.0c01140
J.C. Yan, T. Gao, Z.Z. Lu, et al., ACS Appl. Mater. Interfaces 13 (2021) 27749–27773. doi: 10.1021/acsami.1c06818
S.Y. Low, J.E. Hill, J. Peccia, Biochem. Biophys. Res. Commun. 378 (2009) 701–705. doi: 10.1016/j.bbrc.2008.11.087
P. Liu, C. Li, R.X. Zhang, et al., Biosens. Bioelectron. 126 (2019) 543–550. doi: 10.1016/j.bios.2018.10.048
J.A. Bosch, C.L. Chen, N. Perrimon, WIRES Dev. Biol. 10 (2021) e392.
S. Han, J. Li, A.Y. Ting, Curr. Opin. Neurobiol. 50 (2018) 17–23. doi: 10.1016/j.conb.2017.10.015
A. Warford, H. Akbar, D. Riberio, Methods 70 (2014) 28–33. doi: 10.1016/j.ymeth.2014.03.001
Q. Zhang, Q.Y. He, J.H. Wang, C.Y. Fu, H.L. Hu, Nat. Protoc. 13 (2018) 118–133. doi: 10.1038/nprot.2017.134
A.M. Maley, P.M. Garden, D.R. Walt, ACS Sens. 5 (2020) 3037–3042. doi: 10.1021/acssensors.0c01661
J.F. Li, S. Han, H.J. Li, Cell 180 (2020) 373–386. doi: 10.3390/w12020373
B.T. Lobingier, R. Huttenhain, K. Eichel, et al., Cell 169 (2017) 350–360. doi: 10.1016/j.cell.2017.03.022
G.B. Mao, Y.X. Ma, G.Q. Wu, et al., Anal. Chem. 93 (2021) 777–783. doi: 10.1021/acs.analchem.0c03078
H.W. Rhee, P. Zou, N.D. Udeshi, et al., Science 339 (2013) 1328–1331. doi: 10.1126/science.1230593
B.W. Liu, S. Salgado, V. Maheshwari, J.W. Liu, Curr. Opin. Colloid Interface Sci. 26 (2016) 41–49. doi: 10.1016/j.cocis.2016.09.001
C.H. Lu, H.H. Yang, C.L. Zhu, X. Chen, G.N. Chen, Angew. Chem. Int. Ed. 48 (2009) 4785–4787. doi: 10.1002/anie.200901479

Figure 1 (A) Procedure of preparation of aptamer-conjugated HRP. (B) Schematic illustration of specific recognition of aptamer to (1, 3)-β-D-glucan and tyramide signal amplification technology for the identification of fungi.

Figure 2 Fluorescence microscope images of yeast incubated with biotin-aptamer at room temperature for different times (0.5 h, 1 h, 1.5 h, 2 h and 2.5 h). DAPI was at 365 nm excitation. Cy3 was at 546 nm excitation (Scale bar: 20 µm).

Figure 3 Fluorescence microscope images of yeast incubated with aptamer-conjugated HRP for 2 h with TSA technology (Scale bar: 20 µm).

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:
 下载:
下载:
