Table 1.
Summary of cases of mitochondria-targeted peptide-based drug delivery systems for cancer therapy.
Citation:
Yuhan Sun, He Zhang, Guangzhao Lu, Huan Wang, Ying Lu, Li Fan. Mitochondria-targeted cancer therapy based on functional peptides[J]. Chinese Chemical Letters,
2023, 34(5): 107817.
doi:
10.1016/j.cclet.2022.107817

Mitochondria-targeted cancer therapy based on functional peptides
English
Mitochondria-targeted cancer therapy based on functional peptides
Abstract:
Mitochondria are critical for tumor growth and metastasis. A number of traditional antitumor drugs have poor water solubility and must penetrate multiple cellular barriers to reach the mitochondria. Because mitochondria have a unique transmembrane potential and an inner membrane with a low permeability, it is difficult for most drugs to enter mitochondria. In recent years, mitochondria-targeted delivery systems that use functional peptides to modify drugs have received increasing attention. Introducing functional peptides can change the original physicochemical properties of drugs and actively target mitochondria. Functional peptide-drug conjugates (PDCs, peptide-drug conjugates) can decompose and release drugs over time or due to certain stimuli in tumors. This preserves the biological activity of the drug while increasing intratumor uptake through the enhanced permeability and retention effect (EPR, the enhanced permeability and retention effect). In this review, we focus on the direction of cancer therapy and review the application of different functional peptides in the mitochondria-targeted tumor treatments reported in recent years.
-
Key words:
- Peptides
- / Mitochondria
- / Cancer therapy
- / Targeting
- / Cell uptake
-
1. Introduction
Mitochondria are subcellular organelles in mammalian cells. They are the energy pumps for cellular adenosine triphosphate (ATP, adenosine triphosphate) production, the main sites for regulating cellular reactive oxygen species (ROS, reactive oxygen species) and redox homeostasis, as well as the centers of intrinsic apoptosis control [1,2]. The abnormalities and dysfunctions of mitochondria lead to a variety of health-threatening diseases, ranging from neurodegenerative diseases [3,4] and cardiovascular diseases [5] to cancers [6]. Most notably, increasing evidence has revealed the profound impacts of mitochondria used for energy production and the biosynthesis of metabolic substrates on cancer growth, proliferation, metastasis, or recurrence [7–16]. To meet the higher metabolic demand due to rapidly proliferating tumor cells, dysregulated and healthy mitochondria typically exhibit several different structures and functions, such as membrane potential, energy production pathway, respiratory rate, and gene mutation [17]. These differences between cancerous and normal mitochondria offer possibilities to achieve effective cancer treatments by selectively killing tumor cells [18]. Therefore, intensive research efforts have been devoted to developing cancer therapies that target mitochondria [6,19,20].
Since the mitochondrial inner membrane is approximately −180 mV, which indicates a high transmembrane potential [21], the use of lipophilic or cationic molecules as mitochondrial targeting agents is widespread. Common molecules are lipophilic cationic triphenylphosphine (TPP, triphenylphosphine) [22], rhodamine 123 [23], and peptides, such as mitochondrial penetrating peptide (MPP, mitochondrial penetrating peptide) [24] and SS (Szeto-Schiller) peptide [25,26]. TPP can reduce the activation energy passing through the biological phospholipid bilayer, thus TPP has the ability to pass through the mitochondrial membrane and aggregate in the mitochondrial matrix [27,28]. Another way to target mitochondria is to use nanoparticles, such as dequalinium micelles (DQAsomes) [29–33], liposomes [34–36], inorganic nanoparticles [37–39], DNA nanostructures [40,41], and polymeric nanomicelles [42–46]. The most significant difference between DQAsomes and other nanoparticles is that they can actively target mitochondria without other targeting groups. DQAsomes, liposome-like vesicles containing dequalinium chloride, can accumulate in mitochondria, destroy mitochondrial membrane potential, and induce apoptosis or necrosis [47]. However, DQAsomes are rarely used in targeted mitochondrial antitumor therapy due to their low ability for endosomal escape and transfection efficiency [48].
Unfortunately, after insoluble drugs are modified, the drug solubility does not change significantly, and it is difficult for TPP to perform modifications directly. Usually, a backbone group needs to be added for the modification. In addition, TPP is cytotoxic at concentrations higher than 10 µmol/L [48]. Some lipophilic cations have poor water solubility and phototoxicity, which in turn lead to poor circulation in the body [49]. In contrast, peptides with biocompatibility and design variations are more advantageous. These peptides have low immunogenicity and can also mediate precise self-assembly. Functional peptides usually have shorter peptide chains [50], which can be modified on small molecule drugs to form peptide-drug conjugates (PDCs). In addition, peptide-based drug delivery systems have the following advantages: a simple preparation process, good targeting, in vivo degradability, and high absorption in vivo [49]. The systems can change the drug's original physicochemical properties and enhance their biological functions; for example, the time spent in blood circulation can be increased and their therapeutic effects can be enhanced. Some PDCs can also self-assemble to form nanoparticles, fibers, or gels under changes in temperature, pH, or noncovalent interactions. In addition to enhancing tumor tissue penetration and targeting mitochondria in tumor cells [50], these functional PDCs have also been shown to inhibit tumor proliferation and metastasis. The advantages of functional peptides have led researchers to design and synthesize various functional peptides for increasing applications in cancer therapy [51]. The purpose of this review was to examine the application of peptides with different functions in targeting tumor mitochondria for therapeutic purposes.
2. Mechanism of mitochondrial targeting
Targeting the mitochondria of tumor cells has been found to be a very promising method of inhibiting tumor proliferation and metastasis [52,53]. At present, some anticancer drugs, such as lonidamine, doxorubicin (DOX), and cisplatin, have the potential ability to act on the mitochondria of tumor cells.
The main targets of tumor cell mitochondria include mitochondrial DNA (mtDNA, mitochondrial DNA), voltage-dependent anion channel (VDAC, voltage-dependent anion channel), Bcl-2 anti-apoptotic protein family, respiratory chain, and transition pore protein subunits related to mitochondrial membrane permeability [54]. Thirteen essential peptides in the oxidative phosphorylation system are encoded by mtDNA, along with 2 rRNAs and 22 tRNAs [55–57]. Compared to nuclear DNA, mtDNA exhibits a higher mutation rate and fewer DNA repair pathways [58,59]. DOX is reported to produce oxygen-free radicals that cause DNA damage [60]. Cisplatin can bind to DNA and induce DNA damage, ultimately inducing mitochondrial apoptosis [61]. The VDAC family is the most abundant pore-forming protein on the outer mitochondrial membrane, of which the multifunctional membrane channel protein VDAC1 functions as an exchange channel for ions and metabolites between the mitochondria and cytoplasm. The increased rate of glycolysis in tumor cells results in significantly higher expression of hexokinase, which is one of the key enzymes of glycolysis. It has been shown that tumor cells mainly express hexokinase Ⅱ (HK-Ⅱ, Hexokinase Ⅱ), and the binding of VDAC1 to HK-Ⅱ can prevent the binding of proapoptotic proteins (such as Bak and Bax) to mitochondria, ultimately inhibiting apoptosis [62–70]. In turn, inhibiting the interaction of VDAC's interaction with HK-Ⅱ can considerably inhibit cellular proliferation and tumor growth [67]. As an effective inhibitor of HK-Ⅱ, lonidamine can effectively trigger the mitochondrial apoptosis pathway by destroying the mitochondrial transmembrane potential [71,72]. Bcl-2 family proteins regulate apoptosis by altering the permeability of mitochondrial outer membranes [21,73,74]. Anti-apoptotic proteins are overexpressed in tumor cells from various types of cancers, such as colon, thyroid, and breast cancers [75], so inhibiting the expression of these proteins can inhibit tumor cell survival [76,77]. At the same time, some drug molecules self-assemble in or near mitochondria to form fibrous structures, which can destroy the integrity of the mitochondrial membrane, leading to mitochondrial dysfunction [78] and achieving the treatment purpose.
3. Peptides for mitochondrial targeting
Mitochondrial-targeting sequences (MTSs) are usually endogenous peptide sequences consisting of 15–55 amino acids. MTSs vary in size and sequence, but they share the following common feature: the amphiphilic α-helical structure that interacts with negatively charged mitochondrial surfaces and is subsequently internalized by mitochondrial membrane transporters [79]. Using MTSs, proteins synthesized by mitochondrial ribosomes can be delivered to mitochondria. However, MTSs have a poor ability to permeate the cell membrane. To improve their membrane permeability, MTSs are often combined with cell-penetrating peptides (CPPs) to deliver small-molecule chemotherapeutic drugs into the mitochondria of tumor cells [59], as shown in Table 1. The linkers (such as disulfide bonds and hydrazone linkers) between peptides and drugs are degraded by glutathione (GSH), which is highly expressed in tumor cells or in a slightly acidic environment. Lin used the mitochondrial-targeting sequences of MLRAALSTARRGPRLSRLL from the mitochondrial-oriented protein aldehyde dehydrogenase (ALDH) that was covalently bound to CPPs containing histidine; in addition, the researchers attached the nonpermeable membrane dye 5-carboxyfluorescein (5-FAM) to verify whether MTS+CPP could successfully enter the mitochondria [79,80]. In the subcellular localization test, the overlap colocalization coefficient (5-FAM and MitoTracker signals) of MTS-modified molecules was 0.001, while the coefficient of MTS+CPP was 0.277. The results showed that MTS+CPP binding had a stronger cellular uptake and mitochondrial targeting than that of MTS alone. pHK is a mitochondrial-targeting peptide (MIASHLLAYFFTELN) derived from the HK-Ⅱ protein terminus and consists of 15 amino acids. pHK weakens the interaction between VDAC1 on the mitochondrial membrane and endogenous HK-Ⅱ and then promote apoptosis. Abiy D Woldetsadik designed the peptide pHK-PAS for the treatment of cervical cancer. The N-terminal end of pHK is covalently coupled to cell-penetrating peptides (PAS), which inhibits mitochondrial respiration and glycolysis, leading to apoptosis [81]. In tumor cells, pHK-PAS exhibits a 3–7-folds higher cellular uptake that of pHK and is much less toxic to noncancerous HEK293 cells with selective cytotoxicity. pHK sequences were also used by Dan Liu to design pal-pKV-pHK for the treatment of lung cancer. The molecules are composed of peptide sequences for cationic cell-penetrating peptides pKV and pHK and are modified with lipid chain pal to increase intracellular delivery and promote apoptosis [54]. Both synthetic pHK-pKV and pal-pHK-pKV localize to mitochondria and trigger mitochondria-mediated apoptosis. Compared to pHK and pHK-pKV, pal-pHK-pKV showed significantly higher cytotoxicity and stronger antiproliferative activity against A549 cells over the entire concentration range. The results showed that pal-pHK-pKV had increased bioactivity in tumor cells in regard to cytotoxicity, cellular uptake, and mitochondrial localization.
Table 1
Aldehyde dehydrogenase-5MTS (ALD5MTS) are MTSs that target the mitochondrial matrix. Klimpel et al. designed an ALD5MTS-modified cytostatic drug, chlorambucil, for a breast tumor treatment. Chlorambucil acts on DNA and binds to the cell-penetrating peptide sC18 to deliver the drug to the mitochondria, leading to mtDNA damage and dysfunction [59]. In addition, based on the N-(2-hydroxypropyl)methacrylamide (HPMA) copolymer, Li et al. designed a mitochondrial targeting system (DOX-R8MTS) to inhibit the spread of breast cancer [82]. HPMA is a nontoxic, biocompatible, water-soluble nanocarrier. The anticancer drug DOX was modified on HPMA, and then linked to a mitochondria-targeting hybrid peptide (R8MTS, consisting of R8 CPP and ALD5MTS) (Fig. 1). This structure significantly enhanced the cellular uptake and mitochondrial targeting ability of the HPMA copolymer. It was demonstrated that P-D-R8MTS had a strong inhibitory effect on tumor growth, and no obvious pulmonary metastatic nodules were found in the mice carrying 4T1. Compared with P-D-R8, which is not modified with MTS, P-D-R8MTS underwent preferable metastasis and invasion suppression. Furthermore, the tumor accumulation of P-D-R8MTS was 2.98 times higher than that of free DOX, which demonstrated that the HPMA copolymer could promote the tumor accumulation and retention of DOX by intratumoral injection.
Figure 1
 Figure 1. Schematic diagram of antitumor effects of ALD5MTS-based mitochondrial targeting peptide. Reproduced with permission [82]. Copyright 2020, Elsevier B.V.
Figure 1. Schematic diagram of antitumor effects of ALD5MTS-based mitochondrial targeting peptide. Reproduced with permission [82]. Copyright 2020, Elsevier B.V.4. Peptides for mitochondrial penetration
MPPs are specific targeting carriers of small molecules localized to the mitochondrial matrix and exhibit improved cellular uptake and mitochondrial membrane penetration. The structure and function of mitochondria in tumor cells are different from those in healthy cells. The mitochondrial membrane potential of tumor cells was higher than that of normal cells [83]. In addition, mitochondria are stimulated by a series of apoptotic signals (e.g., Bax and Bak) in tumor cells, leading to increased permeability of the outer mitochondrial membrane [84]. Thus, MPPs preferentially target tumor cell mitochondria rather than healthy cells [85,86]. The characteristic by which MPPs pass through cell and mitochondrial membranes is attributed to their positive charge and hydrophobic amino acids [87,88]. Cations promote charge-driven intracellular uptake, and with the help of lipophilic amino acid sequences, MPPs can better pass through the hydrophobic inner mitochondrial membrane (IMM). MPPs can achieve both high cell permeability and mitochondrial localization, as shown in Table 2. Lei and Kelley used disulfide bonds linking MPPs and luminespib for the treatment of leukemia [24]. Luminespib is a mitochondrial heat shock protein HSP90 inhibitor that induces cell cycle arrest and apoptosis. Disulfide bonds were cleaved by GSH, which is highly expressed in tumor cells, and at the same time, luminespib was delivered into the mitochondrial matrix to induce apoptosis. Burke's group combined Ru(Ⅱ) complexes with MPPs to accurately target mtDNA in living cells. Ru(Ⅱ) luminescence, a cell imaging probe, acted as a light switch when it bound with dipyridyl phenazine (dppz) to form the Ru(Ⅱ) complex. The probe was photoswitched on when bound to mtDNA and photoexcited in hydrophilic media. Binding of Ru(Ⅱ) to MPP can be used as a fluorescent reporter molecule for targeting cellular mtDNA [56].
Table 2
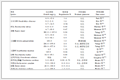 Table 2. Summary of cases of mitochondria-penetrating peptide-based drug delivery systems for cancer therapy.
Table 2. Summary of cases of mitochondria-penetrating peptide-based drug delivery systems for cancer therapy. DownLoad:
CSV
DownLoad:
CSV

To combat tumor resistance and metastasis, Deng et al. modified α-cyclodextrin (α-CD)-based doxorubicin, NO prodrugs, and dimethylmaleic anhydride (DA) onto polyethylene glycol (PEG)-conjugated MPPs (KLAKLAK)2CGKRK to obtain α-CD-DOX-NO-DA nanoparticles (Fig. 2A). Adding PEG to nanoparticles increased their circulation time in the blood and cellular uptake capacity. The release of NO induced mitochondrial dysfunction and effectively overcame tumor drug resistance [89]. In an in vivo antitumor assay, the tumor growth inhibition ratio (TIR) of α-CD-DOX-NO-DA nanoparticles at day 24 was 86.8%, which was much higher than that of free DOX (11.4%). In a study on the inhibition of in situ drug-resistant tumors, free DOX did not inhibit tumor growth, while α-CD-DOX-NO-DA nanoparticles almost completely inhibited tumor growth. In addition, the weight change of mice during the treatment was negligible, indicating that α-CD-DOX-NO-DA nanoparticles had low systemic toxicity.
Figure 2
 Figure 2. (A) Schematic structure and action of drug delivery system based on KLAK peptide with mitochondrial penetrating function. Reproduced with permission [89]. Copyright 2020, WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. (B) Schematic structure and action of drug delivery system based on FxrFxK peptide with mitochondrial penetrating function. Reproduced with permission [90]. Copyright 2021, Elsevier B.V.
Figure 2. (A) Schematic structure and action of drug delivery system based on KLAK peptide with mitochondrial penetrating function. Reproduced with permission [89]. Copyright 2020, WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. (B) Schematic structure and action of drug delivery system based on FxrFxK peptide with mitochondrial penetrating function. Reproduced with permission [90]. Copyright 2021, Elsevier B.V.Yang et al. designed the MPP-modified HPMA polymer HPMA-MPP-DOX (PM) and HPMA-DOX copolymer (PN) (Fig. 2B), which were co-delivered to inhibit breast cancer metastasis [90]. PM damaged mitochondria to promote apoptosis and inhibit tumor metastasis, while PN disrupted nuclei to inhibit cell proliferation and promote apoptosis. Both in vivo and in vitro experiments showed that the mitochondrial targeting of PM significantly inhibited tumor metastasis and decreased the number of metastatic lung nodules by 67%. The combination of PM and PN complemented each other and synergistically inhibited tumor growth and metastasis. The results showed that this method almost eliminated tumor metastasis, and no obvious pulmonary nodules were observed. Czupiel designed a combination of cationic cell-penetrating peptides (H8R8) and a fatty acid chain (vitamin E succinate or stearyl) to treat multidrug-resistant breast cancer. The structural properties of these peptides are similar to those of MPPs [91], which kill tumor cells by disrupting mitochondria, reducing P-glycoprotein (P-gp) efflux, and inducing ROS production.
5. Peptides for mitochondrial destruction
Mitochondrial destroying peptides are a kind of peptide that utilize self-assembly characteristics and combine with nanotargeted ligands to achieve in vivo morphological transformation and ultimately induce cell dysfunction. Mitochondrial destroying peptides can accelerate the formation of nanofibers under special conditions (such as enzymes and photothermal agents) [92–96]. Due to the increased mitochondrial membrane permeability and potential of tumor cells, the targeted drug delivery systems mediated by mitochondrial destruction peptides tend to aggregate in tumor cells and reach critical aggregation concentrations more readily than normal cells. Self-assembled systems play an important role in physiological functions and have great potential for applications in areas such as biomedicine and drug delivery. Self-assembled peptides are formed through various noncovalent interactions, such as hydrogen bonds, π-π superposition, electrostatic interactions, hydrophobic interactions, and van der Waals interactions [41,42]. In organisms, self-assembly peptide has good biocompatibility and biodegradability and a controlled assembly function, which enables aggregation and retention in the focal region.
5.1 Enzyme-instructed intracellular self-assembly (EISA)
EISA is an effective method to induce cellular dysfunction. Drug precursors undergo molecular transformations in the presence of enzymes, changing from soluble hydrophilic units into self-assembling ordered structures that are located intracellularly or extracellularly [94]. Self-assembly of peptides only occurs if the molecular concentration is higher than the critical aggregation concentration (CAC). For mitochondrial self-assembly mediated by mitochondrial targeting peptides, molecules must be enriched in mitochondria. The concentration should be higher than the CAC in mitochondria and lower than the CAC in the cytoplasm. Specific enzymes and excess ROS can be used to trigger the self-assembly process around or on the surface of mitochondria [52]. Thus, the dual action of highly expressed enzymes (e.g., alkaline phosphatase (ALP), GSH) and mitochondrial targeting peptides in tumor cells enables tumor cell mitochondria to be better targeted. However, the shortcomings of EISA are also obvious. The initial concentration of peptides used for self-assembly is higher than that of the critical concentration to form nanofibers [97]. In addition, the use of enzymes is usually limited to certain cell types and cannot be generalized to all tumor cell types. Huaimin Wang designed mitochondrial-targeting compounds (L-1P or D-1P) for the treatment of osteosarcoma [98], as shown in Table 3. The molecules consist of the following parts: (1) the self-assembling backbone tetrapeptide, FFYK; (2) the mitochondrial target head, TPP; (3) an enzymatic triggers (tyrosine phosphate lipids (pY) as a substrate of ALP); and (4) the reporter of molecular self-assembly, an environment-sensitive fluorophore (4-nitro-2,1,3-benzoxadiazole, NBD). The molecules first formed oligomers at certain concentrations and subsequently interacted to form nanoscale assemblies. In addition, due to the rapid dephosphorylation of drug precursors catalyzed by ALP, which are overexpressed on the surface of tumor cells, the oligomers underwent self-assembly and formed nanoscale assemblies on the surface of tumor cells. Through self-assembly, the oligomers formed nanoscale assemblies on the surface of tumor cells, which then entered the tumor cells via endocytosis. Through the targeting effect of TPP, the assemblies accumulate in mitochondria, causing mitochondrial dysfunction and cytochrome C release, ultimately resulting in cell death.
Table 3
Table 3. Summary of cases of mitochondrial-destruction peptide-based drug delivery systems for cancer therapy.DownLoad:
CSV

Jingbo Wang's team prepared a mitochondrial-targeted drug delivery system called DOX@MT-NPs [99]. The molecules consist of the following parts (Fig. 3A): (1) the anticancer drug DOX; (2) the mitochondrial targeting ligand TPP; (3) the peptide-linked block copolymer PEG-b-PPDSEMA; and (4) the self-assembled peptide FFC. These molecules first formed micelles in phosphate buffer solution (PBS). After entering tumor cells in the micelle state, the disulfide bonds between FFC(TPP) and PEG-b-PPDSEMA were enzymatically cleaved by GSH with high expression, resulting in the release of DOX and FFC(TPP). FFC(TPP) gathered and self-assembled into nanofibers after entering mitochondria, which eventually led to mitochondrial dysfunction. DOX acted on DNA and synergistically induced tumor cell death.
Figure 3
 Figure 3. (A) Schematic diagram of mitochondria-targeted drug delivery system DOX@MT-NPs based on enzyme-directed intracellular self-assembly. Reproduced with permission [99]. Copyright 2022, American Chemical Society. (B) Schematic diagram of mitochondria-targeted drug delivery system PKK-S-PEG based on the photothermal promotion of morphological transformation strategy. Reproduced with permission [103,104]. Copyright 2020, American Chemical Society.
Figure 3. (A) Schematic diagram of mitochondria-targeted drug delivery system DOX@MT-NPs based on enzyme-directed intracellular self-assembly. Reproduced with permission [99]. Copyright 2022, American Chemical Society. (B) Schematic diagram of mitochondria-targeted drug delivery system PKK-S-PEG based on the photothermal promotion of morphological transformation strategy. Reproduced with permission [103,104]. Copyright 2020, American Chemical Society.π-π stacking can drive peptide self-assembly, especially for π-coupled peptides (e.g., aromatic peptides) [100]. Phe-Phe (FF, part of β-like amyloid, Aβ19–20) dipeptides, which are used as supramolecular building blocks, can self-assemble to form nanotubes with unique mechanical and electrical properties [35].
Saha's team designed and constructed two lipophilic cationic fluorescent peptide molecules (Cy-3-TPP/FF and Cy-5-TPP/FF) for the treatment of cervical epithelioid carcinoma and lung cancer [101]. The molecules consist of unsymmetrical visible Cy-3 or near-infrared (NIR) Cy-5 chromophores, TPP, and FF dipeptides that can self-assemble to form nanotubes. Cy-3-TPP/FF and Cy-5-TPP/FF molecules were self-assembled in solution to form visible and near-infrared nanotubes, respectively. Modifying TPP groups could effectively transport supramolecular peptide building blocks with a near-infrared imaging function into the mitochondria. These molecules can selectively accumulate in negatively charged mitochondria in tumor cells and form supramolecular nanotubes after reaching CAC. The noncovalent interaction between nanotubes and malignant mitochondrial proteins may be beneficial for inducing early apoptosis.
5.2 Photothermal promotion of the morphological transformation strategy
It was found that peptide assemblies generally aggregate slowly and that increasing the temperature reduces the Gibbs free energy, thereby improving the self-assembly rate [102]. Increased assembly rates encourage the accumulation of drug molecules at the tumor site. Through the photothermal-promoted morphology transformation (PMT) strategy, which involves adding photothermal molecules into PDCs, the local temperature is increased, accelerating the speed of the chemical assembly process and regulating the assembly rate of nanofibers in vivo, ultimately improving the biological properties of drug molecules. Compared to the spontaneous morphology transformation (SMT) strategy, the PMT strategy has a 4-fold increase in the transformation rate. In addition, due to the increased assembly rate, the drug accumulation in tumors under light irradiation is twice as rapid as that in the absence of light irradiation. At the same time, the PMT strategy can induce enhanced accumulation of nanomaterials, which maximizes drug retention at the tumor site.
Compared with nanoparticles, nanofibers have higher Hill coefficients, stronger multivalent interactions, and higher binding affinity with mitochondrial membranes. The conversion of nanoparticles near mitochondria into nanofibers can greatly improve the mitochondrial targeting ability and promote tumor cell apoptosis. Zhang et al. designed and obtained polymer-peptide conjugates (PPCs) PKK-S-PEG for the treatment of breast cancer, and the molecules consist of the following parts [103,104]: (1) peptides that target and disrupt mitochondria [KLAKLAK]2; (2) hydrogen-bonding peptides KLVFF; (3) a hydrophilic PEG fragment that binds disulfide bonds; and (4) a photothermal molecule (purpurin-18, P18). When hydrophilic PKK-S-PEG nanoparticles entered tumor cells through the endocytic pathway, the disulfide bonds between PEG and KLVFF were cleaved by the overexpressed GSH in the tumor cell cytoplasm (Fig. 3B). Then the nanoparticles are transformed into nanofibers (NFs) by the hydrogen bonding of KLVFF. The photothermal effect of P18 was stimulated by NIR radiation, which led to a controlled acceleration of morphological transformation and intensified the induction of apoptosis in tumor cells.
6. Peptides for mitochondrial responsiveness
The addition of a tumor-targeting fraction and antifouling segment to drug carriers is an effective strategy to form nanoplatforms for cancer therapy, which can both prolong blood circulation time and enhance the level of drug delivery within tumor cells [105,106]. Since the important characteristics of the tumor microenvironment include the significantly low pH, low oxygen content, high pressure, and dysregulated enzymatic activity, it can deactivate the performance of antifouling segments and increase the accumulation of nanomaterials within the tumor. The peptide sequence GFLG acts as an environmentally-responsive peptide, which can be cleaved by cathepsin-B (CB, a native lysosomal protease) overexpressed in tumor cells. GFLG formed switchable multifunctional peptides between an outer antifouling fragment and an inner mitochondrial targeting peptide. These multifunctional peptides exhibit a long-term blood circulation time and mitochondrial localization ability.
Xiaokang Jin used this method to design AuNR@FrFK-GFLG-EK for the treatment of breast cancer. The molecules consist of the following parts [105]: (1) the mitochondrial targeting peptide segments FrFK; (2) nanostructured protective group amphipathic peptide sequences EK (glutamic acid, E; lysine, K); (3) CB cleavage site GFLG peptides, which is externally connected to EK peptide and internally connected to FrFK; and (4) gold nanorods, which are used as photothermal agents (PA). The peptides (FrFK-GFLG-EK) were modified on gold nanorods to target mitochondria. Once the molecules reached the tumor site via the enhanced permeability and retention effect (EPR), the environmentally responsive peptide GFLG was cleaved. The external antifouling fragments were separated from the internal mitochondria targeting site, allowing the gold nanorods to target mitochondria and destroy mitochondria in combination with photothermal therapy to achieve therapeutic effects.
7. Conclusion and outlook
In this review, we described mitochondria-targeted cancer therapy strategies based on functional peptides and some promising examples of the rational design of drugs or nanoparticles modified by functional peptides in targeting mitochondria for cancer therapy. Currently, the applications of these functional peptide-based targeted mitochondrial drugs for cancer therapy mainly include the delivery of anticancer drugs or nanoparticles to mitochondria, destruction of mitochondria by mitochondria-cytotoxic peptides or peptide assemblies, and combination with chemotherapy or PMT. Owing to its several advantages, including excellent mitochondria targeting, biocompatibility, and inclusivity for multiple biological and physicochemical activities, the mitochondria-targeted peptide achieves accurate deliveries from the levels of the tissue through cells to cellular organelles [52].
However, several challenges still exist for the further development of mitochondria-targeted peptides. First, rationally designing functional peptides to form robust nanostructures in vivo is still challenging [107]. Second, PDCs have been reported to be mostly responsive to single-factor stimuli. Third, while some studies have demonstrated the biocompatibility of functional peptides, their degradation rate and long-term safety are still debatable. Fourth, although the combination of drugs with mitochondria-targeted functional peptides can enhance tumor cell targeting, it does not completely prevent damage to normal cells.
Two PDCs have been approved by the FDA for clinical trials, including PEPAXTO for the treatment of recurrent bone marrow cancer [108], and LUTATHERA for the treatment of somatostatin receptor-positive gastrointestinal and pancreatic neurosecretory tumors [109]. More than 20 PDCs are being evaluated in clinical trials for cancer therapy [110]. Only a few mitochondria-targeted peptides, such as SS peptides, have successfully advanced to the stage of clinical development for age-related macular degeneration and primary mitochondrial myopathy treatment [111,112]. To date, mitochondria-targeted delivery systems using functional peptides for cancer therapy are still in the primary stage, and many promising results but few clinical reports have been obtained. Despite these challenges, we believe that a more precise mitochondria-targeted peptide-based method with more efficiency and minimal toxicity will be developed for the treatment of cancer.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
-
-
[1]
A.J. Roger, S.A. Muñoz-Gómez, R. Kamikawa, Curr. Biol. 27(2017) R1177-R1192. doi: 10.1016/j.cub.2017.09.015
-
[2]
P. Ning, W. Wang, M. Chen, Y. Feng, X. Meng, Chin. Chem. Lett. 28(2017) 1943–1951. doi: 10.1016/j.cclet.2017.09.026
-
[3]
M.J. Devine, J.T. Kittler, Nat. Rev. Neurosci. 19(2018) 63–80.
-
[4]
J. Döhla, E. Kuuluvainen, N. Gebert, et al., Nat. Cell Biol. 24(2022) 148–154. doi: 10.1038/s41556-021-00837-0
-
[5]
G. Siasos, V. Tsigkou, M. Kosmopoulos, et al., Ann. Transl. Med. 6(2018) 256–278. doi: 10.21037/atm.2018.06.21
-
[6]
S. Fulda, L. Galluzzi, G. Kroemer, Nat. Rev. Drug Discov. 9(2010) 447–464. doi: 10.1038/nrd3137
-
[7]
T. Hu, Z. Qin, C. Shen, H.L. Gong, Z.Y. He, Front. Bioeng. Biotechnol. 9(2021) 786621. doi: 10.3389/fbioe.2021.786621
-
[8]
P.J. Burke, Trends Cancer 3(2017) 857–870. doi: 10.1016/j.trecan.2017.10.006
-
[9]
X.S. Hou, H.S. Wang, B.P. Mugaka, G.J. Yang, Y. Ding, Biomater. Sci. 6(2018) 2786–2797. doi: 10.1039/C8BM00673C
-
[10]
Y. Huang, T. Wang, Q. Tan, et al., Int. J. Nanomed. 16(2021) 4117–4146. doi: 10.2147/IJN.S315368
-
[11]
B. Kalyanaraman, G. Cheng, M. Hardy, et al., Redox Biol. 14(2018) 316–327. doi: 10.1016/j.redox.2017.09.020
-
[12]
K. Klein, K. He, A.I. Younes, et al., Front. Immunol. 11(2020) 573326. doi: 10.3389/fimmu.2020.573326
-
[13]
S. Missiroli, M. Perrone, I. Genovese, P. Pinton, C. Giorgi, eBioMedicine 59(2020) 102943. doi: 10.1016/j.ebiom.2020.102943
-
[14]
P.E. Porporato, N. Filigheddu, J.M.B. Pedro, G. Kroemer, L. Galluzzi, Cell Res. 28(2018) 265–280. doi: 10.1038/cr.2017.155
-
[15]
S. Srinivasan, M. Guha, A. Kashina, N.G. Avadhani, Biochim. Biophys. Acta Bioenerg. 1858(2017) 602–614. doi: 10.1016/j.bbabio.2017.01.004
-
[16]
W.X. Zong, J.D. Rabinowitz, E. White, Mol. Cell 61(2016) 667–676. doi: 10.1016/j.molcel.2016.02.011
-
[17]
M.T. Jeena, S. Kim, S. Jin, J.H. Ryu, Cancers 12(2019) 26–46. doi: 10.3390/cancers12010026
-
[18]
H. Qi, Y. Xu, P. Hu, C. Yao, D. Yang, Chin. Chem. Lett. 33(2022) 1131–1140. doi: 10.1016/j.cclet.2021.09.026
-
[19]
K.G. Roth, I. Mambetsariev, P. Kulkarni, R. Salgia, Trends Mol. Med. 26(2020) 119–134.
-
[20]
Y. Sun, A. Zhan, S. Zhou, et al., Chin. Chem. Lett. 30(2019) 1435–1439. doi: 10.1016/j.cclet.2019.05.001
-
[21]
Y. Gao, H. Tong, J. Li, et al., Front. Bioeng. Biotechnol. 9(2021) 720508. doi: 10.3389/fbioe.2021.720508
-
[22]
Z. Ma, Y. Zhang, J. Zhang, et al., ACS Appl. Mater. Interfaces 12(2020) 39434–39443. doi: 10.1021/acsami.0c11469
-
[23]
R.C. Scaduto Jr., L.W. Grotyohann, Biophys. J. 76(1999) 469–477. doi: 10.1016/S0006-3495(99)77214-0
-
[24]
E.K. Lei, S.O. Kelley, J. Am. Chem. Soc. 139(2017) 9455–9458. doi: 10.1021/jacs.7b04415
-
[25]
W. Mitchell, E.A. Ng, J.D. Tamucci, et al., J. Biol. Chem. 295(2020) 7452–7469. doi: 10.1074/jbc.RA119.012094
-
[26]
H.H. Szeto, AAPS J. 8(2006) E277–E283. doi: 10.1007/BF02854898
-
[27]
M.P. Murphy, R.C. Hartley, Nat. Rev. Drug Discov. 17(2018) 865–886. doi: 10.1038/nrd.2018.174
-
[28]
J. Zielonka, J. Joseph, A. Sikora, et al., Chem. Rev. 117(2017) 10043–10120. doi: 10.1021/acs.chemrev.7b00042
-
[29]
Y. Bae, M.K. Jung, S.J. Song, et al., Mitochondrion 37(2017) 27–40. doi: 10.1016/j.mito.2017.06.005
-
[30]
C. Bailly, Biochem. Pharmacol. 186(2021) 114467. doi: 10.1016/j.bcp.2021.114467
-
[31]
S. Hong, X. Zhang, R.J. Lake, et al., Chem. Sci. 11(2019) 713–720.
-
[32]
M. Shi, J. Zhang, X. Li, et al., Int. J. Nanomed. 13(2018) 4209–4226. doi: 10.2147/IJN.S163858
-
[33]
V. Weissig, M. Lozoya, N. Yu, G.G.M. D'Souza, Methods Mol. Biol. 2275(2021) 13–25.
-
[34]
Y. Feng, G. Qin, S. Chang, et al., Int. J. Nanomed. 16(2021) 3073–3089. doi: 10.2147/IJN.S297716
-
[35]
D.A. Kuznetsova, G.A. Gaynanova, L.A. Vasileva, et al., J. Mater. Chem. B 7(2019) 7351–7362. doi: 10.1039/C9TB01853K
-
[36]
C. Yue, Y. Yang, J. Song, et al., Nanoscale 9(2017) 11103–11118. doi: 10.1039/C7NR02193C
-
[37]
P. Ning, L. Huang, Y. Bao, et al., Bioconjug. Chem. 31(2020) 2719–2725. doi: 10.1021/acs.bioconjchem.0c00518
-
[38]
O. Oladimeji, J. Akinyelu, M. Singh, J. Biomed. Nanotechnol. 16(2020) 853–866. doi: 10.1166/jbn.2020.2930
-
[39]
W. Wang, J. Liu, W. Feng, et al., Biomater. Sci. 7(2019) 1052–1063. doi: 10.1039/C8BM01414K
-
[40]
L. Luo, M. Wang, Y. Zhou, et al., Anal. Chem. 93(2021) 6715–6722. doi: 10.1021/acs.analchem.1c00176
-
[41]
J. Yan, J. Chen, N. Zhang, et al., J. Mater. Chem. B 8(2020) 492–503. doi: 10.1039/C9TB02266J
-
[42]
L.H. Dian, Y.J. Hu, J.Y. Lin, et al., Int. J. Nanomed. 13(2018) 719–731. doi: 10.2147/IJN.S150140
-
[43]
Z. Fan, B. Jiang, D. Shi, et al., Int. J. Pharm. 594(2021) 120184. doi: 10.1016/j.ijpharm.2020.120184
-
[44]
H. Wang, W. Shi, D. Zeng, et al., J. Nanobiotechnol. 19(2021) 152. doi: 10.1186/s12951-021-00895-4
-
[45]
H. Wang, F. Zhang, H. Wen, et al., J. Nanobiotechnol. 18(2020) 8–27. doi: 10.1186/s12951-019-0562-3
-
[46]
Y. Zhang, C. Zhang, J. Chen, et al., ACS Appl. Mater. Interfaces 9(2017) 25152–25163. doi: 10.1021/acsami.7b07219
-
[47]
S. Tang, Z. Davoudi, G. Wang, et al., Chem. Soc. Rev. 50(2021) 12679–12701. doi: 10.1039/D1CS00029B
-
[48]
J. Xu, W. Du, Y. Zhao, et al., Acta Pharm. Sin. B 12(2022) 2778–2789. doi: 10.1016/j.apsb.2022.03.001
-
[49]
L. Huang, Z. Sun, Q. Shen, et al., Chin. Chem. Lett. 33(2022) 4146–4156. doi: 10.1016/j.cclet.2022.02.047
-
[50]
Y. Wang, A.G. Cheetham, G. Angacian, et al., Adv. Drug Deliv. Rev. 110-111(2017) 112–126. doi: 10.1016/j.addr.2016.06.015
-
[51]
N. Mehrotra, S. Kharbanda, H. Singh, Nanomedicine (Lond) 15(2020) 2201–2217. doi: 10.2217/nnm-2020-0220
-
[52]
Z. Luo, Y. Gao, Z. Duan, Y. Yi, H. Wang, Front. Bioeng. Biotechnol. 9(2021) 782234. doi: 10.3389/fbioe.2021.782234
-
[53]
X. Li, W. Zhang, Q. Cao, et al., Cell Death Discov. 6(2020) 80–94.
-
[54]
D. Liu, A. Angelova, J. Liu, et al., J. Mater. Chem. B 7(2019) 4706–4716. doi: 10.1039/C9TB00629J
-
[55]
R.W. Taylor, D.M. Turnbull, Nat. Rev. Genet. 6(2005) 389–402. doi: 10.1038/nrg1606
-
[56]
C.S. Burke, A. Byrne, T.E. Keyes, Angew. Chem. Int. Ed. 57(2018) 12420–12424. doi: 10.1002/anie.201806002
-
[57]
P.F. Chinnery, G. Hudson, Br. Med. Bull. 106(2013) 135–159. doi: 10.1093/bmb/ldt017
-
[58]
A.W. El-Hattab, W.J. Craigen, F. Scaglia, Biochim. Biophys. Acta Mol. Basis Dis. 1863(2017) 1539–1555. doi: 10.1016/j.bbadis.2017.02.017
-
[59]
A. Klimpel, I. Neundorf, J. Control. Release 291(2018) 147–156. doi: 10.1016/j.jconrel.2018.10.029
-
[60]
H. Li, W. Xu, F. Li, et al., Drug Deliv. 29(2022) 192–202. doi: 10.1080/10717544.2021.2023697
-
[61]
M. Kleih, K. Bopple, M. Dong, et al., Cell Death Dis. 10(2019) 851–863. doi: 10.1038/s41419-019-2081-4
-
[62]
G. Calmettes, B. Ribalet, S. John, et al., J. Mol. Cell Cardiol. 78(2015) 107–115. doi: 10.1016/j.yjmcc.2014.09.020
-
[63]
G.S. Krasnov, A.A. Dmitriev, V.A. Lakunina, A.A. Kirpiy, A.V. Kudryavtseva, Expert Opin. Ther. Targets 17(2013) 1221–1233. doi: 10.1517/14728222.2013.833607
-
[64]
S.P. Mathupala, Y.H. Ko, P.L. Pedersen, Oncogene 25(2006) 4777–4786. doi: 10.1038/sj.onc.1209603
-
[65]
J.G. Pastorino, J.B. Hoek, Curr. Med. Chem. 10(2003) 1535–1551. doi: 10.2174/0929867033457269
-
[66]
J.G. Pastorino, J.B. Hoek, J. Bioenerg. Biomembr. 40(2008) 171–182. doi: 10.1007/s10863-008-9148-8
-
[67]
S. Reina, V. De Pinto, Curr. Med. Chem. 24(2017) 4447–4469.
-
[68]
V. Shoshan-Barmatz, D. Ben-Hail, L. Admoni, Y. Krelin, S.S. Tripathi, Biochim. Biophys. Acta 1848(2015) 2547–2575. doi: 10.1016/j.bbamem.2014.10.040
-
[69]
R.J. Winquist, V.K. Gribkoff, Biochem. Pharmacol. 177(2020) 113995. doi: 10.1016/j.bcp.2020.113995
-
[70]
A. Magri, A. Messina, Curr. Med. Chem. 24(2017) 4470–4487.
-
[71]
A.G. Assanhou, W. Li, L. Zhang, et al., Biomaterials 73(2015) 284–295. doi: 10.1016/j.biomaterials.2015.09.022
-
[72]
Y. Liu, X. Zhang, M. Zhou, et al., ACS Appl. Mater. Interfaces 9(2017) 43498–43507. doi: 10.1021/acsami.7b14577
-
[73]
S. Zhang, A. Long, A.J. Link, ACS Synth. Biol. 1(2012) 89–98. doi: 10.1021/sb200002m
-
[74]
T.O. Jose-Luis Diaz, W. Horne, M. McConnell, et al., J. Biol. Chem. 17(1997) 11350–11355.
-
[75]
F. Llambi, D.R. Green, Curr. Opin. Genet. Dev. 21(2011) 12–20. doi: 10.1016/j.gde.2010.12.001
-
[76]
A. Shteinfer-Kuzmine, Z. Amsalem, T. Arif, A. Zooravlov, V. Shoshan-Barmatz, Mol. Oncol. 12(2018) 1077–1103. doi: 10.1002/1878-0261.12313
-
[77]
M. Li, Y. Song, N. Song, et al., Nano. Lett. 21(2021) 5730–5737. doi: 10.1021/acs.nanolett.1c01469
-
[78]
M.T. Jeena, L. Palanikumar, E.M. Go, et al., Nat. Commun. 8(2017) 26. doi: 10.1038/s41467-017-00047-z
-
[79]
S. Kim, H.Y. Nam, J. Lee, J. Seo, Biochemistry 59(2020) 270–284. doi: 10.1021/acs.biochem.9b00857
-
[80]
R. Lin, P. Zhang, A.G. Cheetham, et al., Bioconjug. Chem. 26(2015) 71–77. doi: 10.1021/bc500408p
-
[81]
A.D. Woldetsadik, M.C. Vogel, W.M. Rabeh, M. Magzoub, FASEB J. 31(2017) 2168–2184. doi: 10.1096/fj.201601173R
-
[82]
Q. Li, J. Yang, C. Chen, et al., J. Control. Release 325(2020) 38–51. doi: 10.1016/j.jconrel.2020.06.010
-
[83]
A. Liu, X. Hou, Y. Ding, Y., Acta Pharm. Sin. 52(2017) 879–887.
-
[84]
V. Gogvadze, S. Orrenius, B. Zhivotovsky, Biochim. Biophys. Acta 1757(2006) 639–647. doi: 10.1016/j.bbabio.2006.03.016
-
[85]
H.Y. Chiu, E.X.Y. Tay, D.S.T. Ong, R. Taneja, Antioxid. Redox Signal. 32(2020) 309–330. doi: 10.1089/ars.2019.7898
-
[86]
J. Wu, J. Li, H. Wang, C.B. Liu, Expert Opin. Drug Deliv. 15(2018) 951–964. doi: 10.1080/17425247.2018.1517750
-
[87]
K.L. Horton, K.M. Stewart, S.B. Fonseca, Q. Guo, S.O. Kelley, Chem. Biol. 15(2008) 375–382. doi: 10.1016/j.chembiol.2008.03.015
-
[88]
T. Zhao, X. Liu, S. Singh, et al., Bioconjug. Chem. 30(2019) 2312–2316. doi: 10.1021/acs.bioconjchem.9b00465
-
[89]
Y. Deng, F. Jia, X. Chen, Q. Jin, J. Ji, Small 16(2020) e2001747. doi: 10.1002/smll.202001747
-
[90]
J. Yang, Q. Li, M. Zhou, et al., Int. J. Pharm. 608(2021) 121077. doi: 10.1016/j.ijpharm.2021.121077
-
[91]
P.P. Czupiel, V. Delplace, M.S. Shoichet, J. Control. Release 305(2019) 210–219. doi: 10.1016/j.jconrel.2019.04.045
-
[92]
M. Abbas, Q. Zou, S. Li, X. Yan, Adv. Mater. 29(2017) 1605021. doi: 10.1002/adma.201605021
-
[93]
Z.H. Wang, L. Chen, W. Li, L. Chen, Y.P. Wang, Mitochondrion 65(2022) 80–87. doi: 10.1016/j.mito.2022.05.002
-
[94]
Z. Zheng, P. Chen, M. Xie, et al., J. Am. Chem. Soc. 138(2016) 11128–11131. doi: 10.1021/jacs.6b06903
-
[95]
J. Zhou, X. Du, C. Berciu, et al., Chem 1(2016) 246–263. doi: 10.1016/j.chempr.2016.07.003
-
[96]
J. Zhou, X. Du, N. Yamagata, B. Xu, J. Am. Chem. Soc. 138(2016) 3813–3823. doi: 10.1021/jacs.5b13541
-
[97]
Q. Yao, Z. Huang, D. Liu, J. Chen, Y. Gao, Adv. Mater. 31(2019) e1804814. doi: 10.1002/adma.201804814
-
[98]
H. Wang, Z. Feng, Y. Wang, et al., J. Am. Chem. Soc. 138(2016) 16046–16055. doi: 10.1021/jacs.6b09783
-
[99]
J. Wang, Q. Zhou, X. Li, D. Dutta, Z. Ge, A.C.S. Macro, Lett. 11(2022) 543–548.
-
[100]
P. Zhu, X. Yan, Y. Su, Y. Yang, J. Li, Chemistry (Easton) 16(2010) 3176–3183.
-
[101]
P.C. Saha, T. Bera, T. Chatterjee, et al., Bioconjug. Chem. 32(2021) 833–841. doi: 10.1021/acs.bioconjchem.1c00106
-
[102]
D. Zhang, G.B. Qi, Y.X. Zhao, et al., Adv. Mater. 27(2015) 6125–6130. doi: 10.1002/adma.201502598
-
[103]
X.H. Zhang, D.B. Cheng, L. Ji, et al., Nano. Lett. 20(2020) 1286–1295. doi: 10.1021/acs.nanolett.9b04752
-
[104]
D.B. Cheng, X.H. Zhang, Y.J. Gao, et al., J. Am. Chem. Soc. 141(2019) 7235–7239. doi: 10.1021/jacs.8b07727
-
[105]
X. Jin, H. Yang, Z. Mao, B. Wang, J. Colloid Interface Sci. 601(2021) 714–726. doi: 10.1016/j.jcis.2021.05.135
-
[106]
L. Wu, B. Lin, H. Yang, et al., Acta Biomater. 86(2019) 363–372. doi: 10.1016/j.actbio.2019.01.026
-
[107]
Z. Feng, H. Wang, F. Wang, et al., Cell Rep. Phys. Sci. 1(2020) 100085. doi: 10.1016/j.xcrp.2020.100085
-
[108]
PEPAXTO Prescribing Information, U.S. FOOD & DRUG ADMINISTRATION, 2020,
https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm?event=BasicSearch.process [2022-06-17]. -
[109]
LUTATHERA Prescribing Information, U.S. FOOD & DRUG ADMINISTRATION, 2020,
https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm?event=BasicSearch.process [2022-06-17]. -
[110]
K. Fosgerau, T. Hoffmann, Drug Discov. Today 20(2015) 122–128. doi: 10.1016/j.drudis.2014.10.003
-
[111]
Safety, Efficacy & Pharmacokinetics of Elamipretide, clinicaltrials. gov, 2020,
http://clinicaltrials.gov/ct2/show/NCT03323749 [2022-06-17]. -
[112]
Safety and Efficacy of Elamipretide Primary Mitochondrial Myopathy, clinicaltrials. gov, 2020,
http://clinicaltrials.gov/ct2/show/NCT03323749 [2022-06-17].
-
[1]
-
Figure 1 Schematic diagram of antitumor effects of ALD5MTS-based mitochondrial targeting peptide. Reproduced with permission [82]. Copyright 2020, Elsevier B.V.
Figure 2 (A) Schematic structure and action of drug delivery system based on KLAK peptide with mitochondrial penetrating function. Reproduced with permission [89]. Copyright 2020, WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. (B) Schematic structure and action of drug delivery system based on FxrFxK peptide with mitochondrial penetrating function. Reproduced with permission [90]. Copyright 2021, Elsevier B.V.
Figure 3 (A) Schematic diagram of mitochondria-targeted drug delivery system DOX@MT-NPs based on enzyme-directed intracellular self-assembly. Reproduced with permission [99]. Copyright 2022, American Chemical Society. (B) Schematic diagram of mitochondria-targeted drug delivery system PKK-S-PEG based on the photothermal promotion of morphological transformation strategy. Reproduced with permission [103,104]. Copyright 2020, American Chemical Society.
Table 1. Summary of cases of mitochondria-targeted peptide-based drug delivery systems for cancer therapy.

 下载: 导出CSV
下载: 导出CSV
Table 2. Summary of cases of mitochondria-penetrating peptide-based drug delivery systems for cancer therapy.
下载: 导出CSV
Table 3. Summary of cases of mitochondrial-destruction peptide-based drug delivery systems for cancer therapy.
下载: 导出CSV
-







 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 18
- 文章访问数: 1369
- HTML全文浏览量: 89
