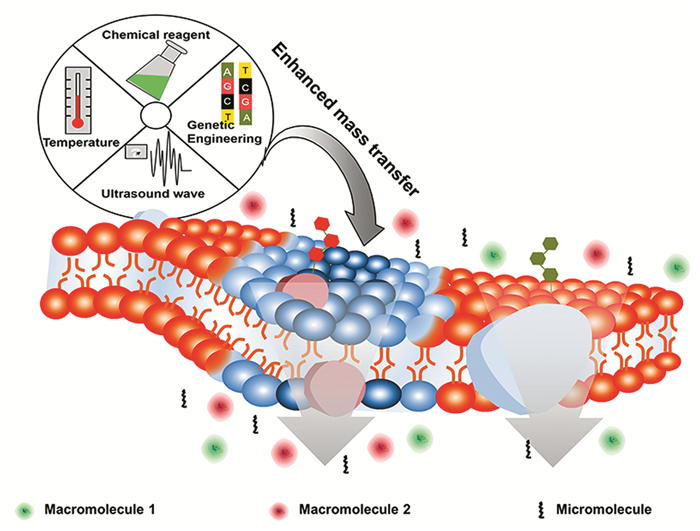
图 1.
细胞透性化技术强化细胞传质
Figure 1.
Cell permeability technology enhances cell mass transfer

过程强化这一概念是在近几十年来化工技术飞速发展而环境资源问题日益严峻的情况下提出的。它旨在减少废料、降低能耗、缩小生产设备尺寸的同时又能提高生产效率,达到事半功倍的效果[1],是当前践行绿色化学发展理念的重要手段。
全细胞催化是指在活细胞或休眠/死细胞的作用下,将底物转化为特定产物的过程[2]。其催化成本低,减少了传统酶催化细胞裂解和蛋白质浓缩等步骤,尤其当涉及到有辅酶因子的复杂转化时,细胞催化的优势尤其明显[3~5]。因此,其在制药[6~10]、工业合成[11~14]、绿色再生能源[15~17]等领域展现出独特的优势和极大的应用前景。然而,细胞的透性屏障作用却成了阻碍细胞催化发展的主要问题,有报道称,游离酶催化速度比细胞催化快10~100倍[18, 19]。以大肠杆菌为例,其细胞膜分为外膜和内膜,外膜面积约为6.7μm2,含有3.5×106个脂多糖分子,占据了菌体表面的四分之三[20]。由于脂多糖分子的两亲性,密集组装的脂多糖无论对于亲水分子还是疏水分子都是不可忽视的屏障。因此,研究者不断尝试用更好的方法来提高细胞膜内外物质的传递速度,减少催化过程中不必要的能耗损失。目前这些方法大致可以分为两种:通过细胞透性化技术和缩小反应规模来实现全细胞催化过程强化。
本文简要分析了为提高细胞催化效率而探索的过程强化方法及各种方法的机理及优缺点,探讨了全细胞催化过程强化的挑战,以期为将来解决此问题的研究提供参考。
细胞透性化技术可以在保持细胞活性和结构的同时,增加细胞膜通透性,从而减少细胞渗透屏障作用,降低胞内外物质传质阻力(见图 1)。其可主要分为化学法、物理法和分子生物学法(见表 1)。
 下载:
导出CSV
下载:
导出CSV
| 方法 | 减少渗透屏障作用机理 | 特点 | |
| 化学法 | 表面活性剂 | 自发插入细胞膜,降低膜边缘张力,膜上打开小孔并变大,使细胞膜不稳定破裂。 | 较为常用,效果显著,价格较低;易造成细胞破碎,难以重复利用。 |
| 有机溶剂 | 与细胞膜进行静电作用,破坏细胞膜亲疏水平衡,改变膜蛋白结构。 | 常用且效果显著;存在使用安全问题;工艺下游溶剂残留问题。 | |
| 共轭低聚物 | 与细胞表面静电吸引插入磷脂双分子层,改变细胞放电能力,影响细胞Zeta电位。 | 对比表面活性剂和有机溶剂细胞毒性较小;对细胞生长代谢具有抑制作用,重复利用率不高。 | |
| 物理法 | 温度 | 高温:膜蛋白变性形成小孔;冻融:菌体内部产生冰晶破坏细胞膜和细胞壁。 | 高温:方法简便易于放大;不耐高温酶不适用;冻融:对细胞内酶活有利;较为繁琐。 |
| 超声 | 对细胞壁和细胞膜结构进行物理破坏。 | 成本较高;不易放大;会造成酶泄露。 | |
| 电渗透 | 改变膜介电常数使代谢物释放。 | 过程可逆,细胞重复利用率高。 | |
| 分子生物学法 | 分子水平上对膜蛋白成分和表达进行改造。 | 对细胞活性几乎无伤害,细胞重复利用率高。 |
化学法是通过表面活性剂、有机溶剂等破坏细胞膜的完整性,使其调控物质传递能力减弱。这种方法效果显著、容易放大,但由于不同细胞的细胞膜结构组分具有多样性,使得此法易导致细胞膜的过度损伤,细胞裂解而无法实现重复利用。
十六烷基三甲基溴化铵(CTAB)是一种常用于改善细胞膜通透性的阳离子季铵盐表面活性剂。Ni等[21]用CTAB处理安大略假丝酵母,透化的酵母细胞活性提高了2倍以上,将(R-2-氯-1-(3-氯苯基)乙醇的催化时间从72h减少至24h。Moon等[22]将包含多聚磷酸激酶的ATP再生系统与CTAB结合使用,把1, 5-戊二胺的产率提高了60%。ATP再生系统能够提高1, 5-戊二胺的产量,而表面活性剂CTAB则可以提高产物跨膜运输效率。Cortez等[23]用Triton X-100(聚乙二醇辛基苯基醚)来处理酵母细胞,改进D-木糖向木糖醇的生物转化效率。与大多数表面活性剂相似,Triton X-100是一个锥形分子,当它越来越多地插入细胞膜中时,膜边缘张力不断降低,膜上小孔不断打开并变大,直到细胞膜最终变得不稳定而破裂。
Kumar等[24]研究了氯仿、乙醚、乙酸乙酯等常用有机溶剂对芳香欧文氏菌通透性及对其中青霉素酰化酶活性的影响。结果表明,甲苯、乙醚等有增加酶活的效果,而丙酮、乙醇则会降低酶活。这与有机溶剂的介电常数和油水分配系数(logP)息息相关,介电常数高于9或亲水性有机溶剂长时间处理细胞,胞内酶的亲水壳层结构会变形,使酶活急剧下降。Zhang等[25]通过戊二醛将细胞与聚乙烯亚胺(PEI)共固定,再用异丙醇对生产(R)-(-)-扁桃酸的重组大肠杆菌进行处理,发现处理过的细胞内腈水解酶活是未处理过的4.6倍,且细胞在循环利用15次后活性未见明显损失。此方法为解决化学透性化技术细胞重复利用率低的问题带来了曙光。此外,他们发现在加入2%的异丙醇时大肠杆菌的催化活性达到最大,继续增加异丙醇用量活性则会下降,这是异丙醇损坏细胞膜从而酶泄露所导致的。
共轭低聚物(COE)是一种具有极大应用前景的合成有机分子,抑菌是其作用之一。它通过与细胞表面的静电吸引自发插入磷脂双分子层中,改变细胞的放电能力,影响细胞的Zeta电位,从而改变细胞膜的通透性[26]。Wang等[27]用4种不同的COE修饰细胞膜,并与常用的表面活性剂CTAB进行对比,发现虽然仍有一定毒性,但其中用COE修饰的细胞比起用CTAB处理有更好的催化速率和产率。
对比起化学法,物理法具有无试剂残留、产物易分离、操作易控制等优势。它主要是通过各种物理作用来改变菌体的细胞被膜结构,从而改善细胞对底物和产物的通透性,提高细胞催化效率。
Isabella等[28]发现,在75℃下处理过的重组大肠杆菌催化藻酸盐的活性提高了4倍。热处理不仅使大肠杆菌膜蛋白变性形成小孔增加了细胞膜通透性,还使一些催化反应不需要的蛋白质失活,减少了副产物的产生。但这对于一些不耐高温的细胞催化反应是不友好的,而冻融法就很好地规避了这个不足。菌体内部在冻融过程中产生的冰晶破坏了细胞膜和细胞壁,提高了细胞通透性[29]。虽然此方法对细胞催化活性是有利的,但其操作繁复,处理时间较长。
相较而言,超声处理法和电渗透法更具优势,它们对细胞膜的通透性处理都是可逆的。细胞超声处理常被用于辅助药物递送,可以促进弱渗透性治疗药物的细胞膜透过率。处理后细胞膜通透性提高,又能在短时间(1~3 h)内恢复[30]。这种“时间窗口”无论对于癌细胞的靶向药物治疗或是细胞催化的催化剂重复利用率都是有利的。Dong等[31]研究了不同频率的超声处理对耻垢分枝杆菌细胞通透性的影响,发现超声处理能破坏细胞壁和细胞膜的物理结构,破坏程度随超声强度的增加而增加。Vaessen等[32]研究了脉冲电场处理过的植物乳杆菌(Plantarum WCFS1)细胞膜通透性的可逆性,发现可逆或不可逆取决于脉冲电场的处理强度,电场强度影响细胞膜上的跨膜电位差,当该电位差大于临界值时,细胞膜上会形成小孔。Tryfona等[33]用脉冲电压处理谷氨酸棒杆菌,使氨基酸的释放提高了几个数量级,这是由于脉冲电压会改变细胞膜的介电常数,诱导代谢物释放。
分子生物学法是在分子水平上对细胞的基因进行敲除、敲入和点突变等,改变细胞原有的遗传特性。近年来,通过基因工程改善细胞膜的传质已经成为了一个热点。
膜蛋白是细胞膜功能的主要承担者,物质转运就是其重要功能之一。理想情况下,我们希望底物无阻滞地被运输到细胞中,生产速率仅由细胞代谢功能决定,一旦产物在细胞内合成并积累到一定水平,就可以被释放并收集。然而,现实情况往往是细胞膜限制了底物与产物的传质,出现细胞内产物抑制,甚至造成催化反应停止。通过分子生物学法对膜蛋白表达进行修饰改造,可以降低底物和产物的传质阻力,提高其传质速率。Julsing等[34]发现,外膜蛋白AlKL的表达将重组大肠杆菌十二烷酸甲酯的催化活性提高了28倍。Jeon等[35]研究了脂肪酸转运蛋白FadL在重组大肠杆菌中的表达水平对催化效率的影响,发现FadL的表达使重组大肠杆菌长链脂肪酸和羟基脂肪酸的最大反应速率增加了5.5倍。Park等[36]发现,来自恶臭假单胞菌的FadL与来自水油海杆菌的细胞色素P450单加氧酶的共表达能加强1-十二烷醇的细胞膜透过率,促进1, 12-十二烷二醇的生物转化。因此,表达长链脂肪酸转运蛋白FadL对于提高疏水性物质的传质具有重要作用。
脂多糖作为大多数革兰氏阴性细菌外膜的主要成分其结构对细胞通透性具有重要影响。脂多糖分子由O抗原、核心多糖与类脂A三部分结构组成,具有两亲性,对于亲水或疏水分子都是一层屏障。通过某些表达脂多糖基因的突变或敲除,改变脂多糖上的磷酸基团结构、脂肪酸链数量、多糖链长度或影响外膜蛋白表达等,将获得高渗透性的细胞。Pagnout等[37]将控制脂多糖合成的waa基因进行点突变,研究突变菌株细胞膜性质的变化,发现突变株ΔwaaC的外膜蛋白显著减少,原本的外膜蛋白被磷脂填充,导致疏水性物质的膜透过率更高;突变株ΔwaaG对疏水性抗生素新霉素和阴离子表面活性剂十二烷基硫酸钠(SDS)具有超敏性。Wang等[38]从大肠杆菌W3110上分别敲除了10个参与脂多糖合成的基因,从而产生了10种大肠杆菌突变株,并研究了各基因缺失对于外膜通透性的影响,发现与原菌株相比,ΔwaaC、ΔwaaF和ΔwaaG突变株的膜渗透率显著提高。显然,脂多糖的核心结构变化对于细胞膜的传质有很大的影响。
微化工技术是过程强化技术的重要组成部分,在微尺度上研究“三传一反”,可以解决高能耗高物耗的问题,实现过程的安全、温和、集成和高效。如图 2所示,微尺度体系的传质传热速率、反应物混合程度以及催化活性位点与反应物之间的接触率对催化过程都具有重要影响[39, 40]。传统的发酵罐细胞催化需要不断搅拌从而使底物与细胞充分接触,而此方法对于细胞活性并不有利,搅拌引起的巨大剪切力会使脆弱的细胞结构破损而失活。在发酵过程进行时,细胞代谢产生的产物与副产物(如乙酸、丙酸等)在发酵罐内不断积累使催化环境pH降低,抑制细胞生长,甚至引起菌体代谢过程改变,影响产物质量。全细胞催化过程受到细胞膜传质阻力的影响较为明显,提高传质扩散效果能较好地提升催化反应过程。微流体反应器当量直径通常小于500μm,较小的体积决定了体系中的雷诺数(Re,一种用来表征流体流动情况的无量纲数)较低[41]。雷诺数低则体系中流体粘度的影响比流体惯性更大,流体处于层流状态,流动稳定有序且层次分明,流体剪切力较小,对细胞活性影响甚微;底物产物的混合均一稳定,产物质量和生产效率的可控性更佳。因而,较小的雷诺数有利于全细胞催化反应的过程强化。雷诺数可用式(1)表示。
|
$ {\rm{Re}} = \frac{{\rho VL}}{\mu } $ |
(1) |

式中,ρ是流体密度系数;μ是动力粘性系数;V是流场的特征速度;L是流体的特征长度。
另外,微反应器更大的比表面积有利于传质和传热,增加了反应物与催化剂接触概率,缩短了传质距离,提高了催化速率[42]。受限的微小空间减少了过程能量的损耗,提高了生产效益;对催化反应条件的精准控制减少了副产物的产生,提高了目标产物的催化选择性。将多个微反应器进行并联或串联就能扩大催化反应规模,有利于其在实验室研究和工业生产中的转换[43]。由此可见,全细胞催化适合在微尺度下进行强化。
微尺度生物催化的主要挑战在于生物催化剂在微流体中的稳定性和重复利用率。微尺度酶催化已有报道[44~46],例如交联并固定在反应器表面[47]、载体固载[48]、微囊包封[49]等方式成功提高了酶的稳定性和利用率。最近,微尺度细胞催化的过程强化已成为研究热点。
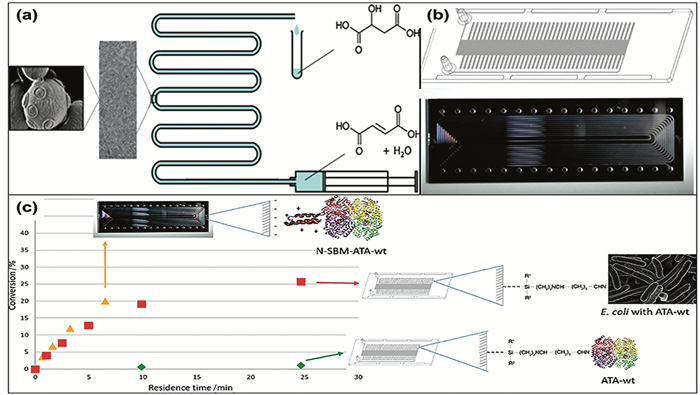
Stojkovic等[50]用3-氨基丙基三乙氧基硅烷(ATPES)和戊二醛处理全氟烷氧基(PFA)微通道(见图 3),APTES用于形成带有裸露氨基的硅烷层,该氨基与戊二醛共价结合,戊二醛再与细胞上的氨基共价结合,由此将细胞沉积在通道壁上。随后,他们用CTAB处理通道壁上的细胞增加细胞通透性,用于连续生产L-苹果酸[51]。这种方法适用于酿酒酵母、枯草芽孢杆菌、恶臭假单胞菌和大肠杆菌等多种细胞,细胞覆盖率最高可达75%,且细胞在4d内都显示良好的稳定性[52]。此方法的优点在于细胞覆盖率较高,然而较多的化学修饰步骤对于细胞活性是不利的[40]。Milozic等[53]用相同的方法修饰微通道,将ATA(氨基转氨酶)、N-SBM-ATA(N-硅胶结合模块标签结合的氨基转氨酶)与重组大肠杆菌分别固定在微反应器内表面进行连续催化反应,对比它们的转化率。结果发现,在微通道上ATA的负载率极低,几乎检测不到产物;N-SBM-ATA具有更大的负载率,但用于连续催化时酶稳定性较差导致转化率不高。相较而言,固载细胞的微反应器具有良好的稳定性,在连续催化4d后,原本固定的细胞仍保留了87%。
水凝胶是对水具有高亲和力的三维网格结构凝胶,生物相容性好,细胞在其中可以长时间保持活性[54, 55]。Menegatti等[56]将载有酵母细胞的海藻酸钠-聚乙烯醇(SA-PVA)共聚物水凝胶引入微反应器中,研究不同交联剂对细胞催化的影响,发现细胞稳定性能维持7d左右。这种方法虽然避免了化学试剂对细胞的损害,但由于水凝胶的传质阻碍,催化效率得不到很好的提高。
生物膜是指物体表面的微生物群体以及微生物形成的絮凝体或凝聚体以及其自身分泌的胞外聚合物,是自然界中微生物存在的主要方式[57]。生物催化反应中,底物或产物的毒性会导致催化剂的不稳定,而生物膜中微生物对于有毒化学物质和物理压力的内在抗性、细胞的自动固定和更新等,使生物膜反应器成为生物催化中的一个热点[58]。Karande等[59]用生长台湾假单胞菌生物膜的微反应器将具有细胞毒性与高挥发性底物环己烷(logP值为1.5)转化为环己醇。为了体系内氧气与环己烷气体的平衡,他们在内径2mm的硅胶管内生长生物膜,将硅胶管置于装有环己烷的瓶中,在硅胶管内分段流入水相与气相,瓶中的环己烷能够透过有机硅膜进入硅胶管中,多余的环己烷不断被水相带走,降低环己烷对细胞活性的不利影响(见图 4(a))。Willrodt等[60]利用恶臭假单胞菌在分段流动生物膜微反应器中将易得的挥发性底物单萜柠檬烯进行生物催化获得高价值的抗癌化合物紫苏酸。恶臭假单胞菌在60mmol/L以上的紫苏酸浓度下生长被完全抑制,分段流动微反应器的应用可以将产物紫苏酸不断带走,减少毒性影响,其紫苏酸产率是补料分批发酵的4倍。Halan等[61]在水-气-有机连续三相分段流动毛细管微反应器中培养台湾假单胞菌生物膜,催化剧毒物质苯乙烯为(S)-苯乙烯氧化物,开发了一种底物有剧毒且难溶于水的生物催化微反应器。先将细胞附着在微通道壁上,连续通过空气和包含葡萄糖的水相培养,6d后开始通入底物苯乙烯, 催化反应刚开始时底物转化率较低而后逐步提高,可能是苯乙烯的接触使细胞结构受损而后逐渐恢复的原因,显示出生物膜微应器的独特优势。
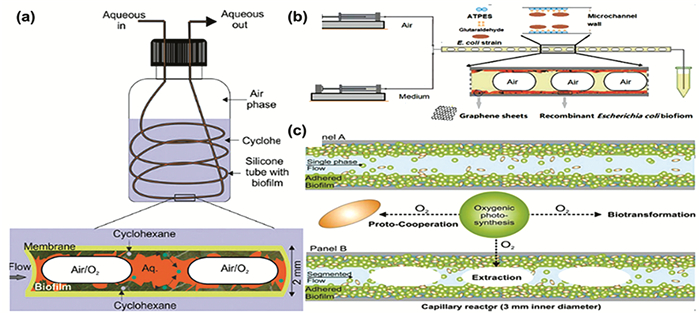
微反应器中细胞的无限增殖难以调控,导致微流体堵塞以及通道内代谢所需氧气的缺乏是生物膜微反应器应用的主要挑战[62]。为了解决这些问题,Zhu等[63]在用ATPES修饰的微通道表面通过共价结合的方式沉积细胞,再分段流入含纳米材料的空气和培养基来调控生物膜的生长,并研究了不同纳米材料对生物膜的生长调节,开发了一种快速有效的生物膜催化剂制备方法(见图 4(b))。Hoschek等[64]设计了一种培养高密度生物膜的方法,在毛细管内表面引入光自养-化学异养双菌株形成生物膜(见图 4(c))。一种菌供氧,一种菌耗氧并进行催化反应生成环己醇,转化率高达98%,细胞稳定性长达1个月。有趣的是,微反应器中的氧气还可以通过向通道中分段通入空气和培养基来调控——多余的氧气会自动混入气泡中。
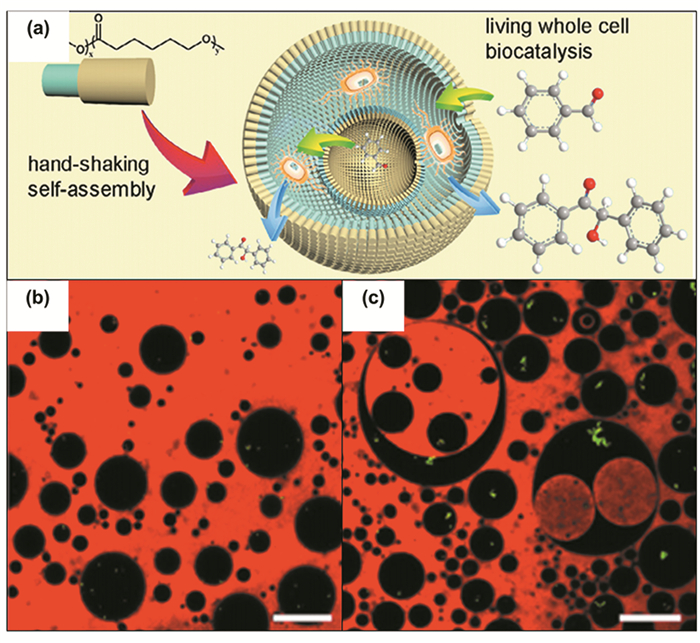
乳液的应用为生物催化开辟了新思路,已经有不少研究利用乳液进行酶催化并取得了很好的结果[47]。最近,有报道将乳液与细胞催化相结合,为细胞建立了具有良好稳定性的微区室[65]。Zhao等[66]通过手动摇晃并改变油水比例获得具有多隔室的乳液,成功包封了活细胞进行生物催化。被包封的细胞回收容易且细胞活性在使用5个循环后仍超过70%(见图 5)。与传统催化体系相比,微区室的超大比表面积强化了底物和产物的传质,将大肠杆菌的催化活性提高了137倍。然而,此方法仍有稳定乳液的乳化剂的传质阻滞、摇晃所得乳液不均一且乳化效率不高等问题。
化工过程强化是我国发展绿色化学,向资源节约型、环境友好型发展模式转变的重要手段。面对当前日益严峻的环境问题,发展生物催化技术已经成为必然趋势。在全细胞催化优势日益突出的今天,亟需发展提高细胞内外物质传递速率的方法。今后的全细胞催化过程强化研究可以从这两个主要方面入手:
(1) 传统的细胞透性化技术需要更多的创新。化学法效果显著且操作简便,主要问题在于如何提高此法的细胞重复率;物理法对细胞活性相对友好但不容易放大,限制了大规模的工业生产;分子生物学法对细胞结构和活性基本无影响,又避免了透性化试剂的处理和工艺下游的分离,相信随着基因工程的发展,对该方法的研究会更加深入。
(2) 近年来兴起的微尺度全细胞催化技术能实现过程的安全、集成、高效、可控,并减少过程的能耗和物耗,具有极大的发展前景。然而在实际应用中,微尺度全细胞催化仍存在成本高、通道堵塞与细胞的分离循环使用等问题。微反应器单元的制作成本较高,如果不能降低成本,微尺度细胞催化的工业生产“数增放大”就无从谈起;细胞作为可以进行生长代谢的独立且有差异的个体,不同于传统的固、液、气微化工反应体系,在“数增放大”后可能出现生长不易控制堵塞通道等问题;如何在实现细胞催化剂高效分离及循环利用的同时又能保证体系较高的传质速率仍然是实现细胞催化从实验室走向工业生产,提高科技成果转化率的关键。随着微尺度过程强化技术在业内认可度越来越高,它在全细胞催化中的应用将会获得长足的发展,为全细胞催化领域带来重大突破。
孙宏伟, 陈建峰.化工进展, 2011, 30(1): 1-15.
郭明, 胡昌华.中国生物工程杂志, 2010, 30(4): 110-115.
Dockrey S A B, Doyon T J, Perkins J C, et al. Chem. Biol. Drug Design, 2019, 93(6): 1207-1213. doi: 10.1111/cbdd.13443
王秋雨, 钦传光, 左小佳, 等.化学通报, 2009, 72(7): 587-593.
赵伟睿, 胡升.中国生物工程杂志, 2014, 34(3): 125-131.
Chen X, Zhou J, Zhang L, et al. Metabol. Eng., 2018, 47(1): 374-382.
Gao Q, Piret J M, Adrio J L, et al. J. Ind. Microbiol. Biotechnol., 2003, 30(3): 190-194. doi: 10.1007/s10295-003-0034-4
Hossain G S, Shin H D, Li J, et al. RSC Adv., 2016, 6(86): 82676-82684. doi: 10.1039/C6RA16507A
Ke C, Yang X, Rao H, et al. Springerplus, 2016, 5(591): 1-8.
Zhu D Q, Zhan X B, Wu J R, et al. Biotechnol. Lett., 2017, 39(1): 55-63. doi: 10.1007/s10529-016-2215-z
Hama S, Tamalampudi S, Suzuki Y, et al. Appl. Microbiol. Biotechnol., 2008, 81(4): 637-645. doi: 10.1007/s00253-008-1689-6
He Y, Chen F, Sun M, et al. Molecules, 2018, 23(3): 691. doi: 10.3390/molecules23030691
Hosche Ak, Buehler B, Schmid A. Biotechnol. Bioeng., 2019, 116(8): 1887-1900. doi: 10.1002/bit.27006
Yamaguchi T, Nuylert A, Ina A, et al. Sci. Rep., 2018, 6: 26998.
Jin G, Bierma T J, Hamaker C G, et al. J. Environ. Sci. Heal. A, 2009, 44(1): 21-28. doi: 10.1080/10934520802515202
Tian K, Li Z. Biochem. Eng. J., 2016, 115(1): 30-37.
Yeom S H. Biotechnol. Bioproc. Eng., 2016, 21(2): 274-282. doi: 10.1007/s12257-016-0046-3
Chen R R. Appl. Microbiol. Biotechnol., 2007, 74(4): 730-738. doi: 10.1007/s00253-006-0811-x
Ni Y, Chen R R. Biotechnol. Bioeng., 2004, 87(6): 804-811. doi: 10.1002/bit.20202
Rietschel E T, Kirikae T, Schade F U, et al. FASEB J., 1994, 8(2): 217-225. doi: 10.1096/fasebj.8.2.8119492
Ni Y, Zhang B H, Sun Z H. Chin. J. Catal., 2012, 33(4): 681-687.
Moon Y M, Yang S Y, Chol T R, et al. Enzyme Microb. Technol., 2019, 127: 58-64. doi: 10.1016/j.enzmictec.2019.04.010
Cortez D V, Mussatto S I, Roberto I C. Appl. Biochem. Biotechnol., 2016, 180(5): 969-979. doi: 10.1007/s12010-016-2146-0
Kumar A, Pundle A. J. Mol. Catal. B, 2009, 57(1-4): 67-71. doi: 10.1016/j.molcatb.2008.06.018
Zhang Z J, Yu H L, Imanaka T, et al. Biochem. Eng. J., 2015, 95(15): 71-77.
Catania C, Thomas A W, Bazan G C. Chem. Sci., 2016, 7(3): 2023-2029. doi: 10.1039/C5SC03046C
Wang B, Fronk S L, Rengert Z D, et al. Chem. Mater., 2018, 30(17): 5836-5840. doi: 10.1021/acs.chemmater.8b02848
Lernia I D, Schiraldi C, Generoso M, et al. Extremophiles, 2002, 6(4): 341-347. doi: 10.1007/s00792-001-0263-2
Cortez D V, Roberto I C. New Biotechnol., 2012, 29(2): 192-198. doi: 10.1016/j.nbt.2011.05.011
Lammertink B, Deckers R, Storm G, et al. Int. J. Pharm., 2015, 482(1-2): 92-98. doi: 10.1016/j.ijpharm.2014.12.013
Dong Y, Su H, Jiang H, et al. Ultrason. Sonochem., 2017, 37: 1-8. doi: 10.1016/j.ultsonch.2016.12.024
Vaessen E M J, Timmermans R A H, Tempelaars M H, et al. Sci. Rep., 2019, 9: 1-11. doi: 10.1038/s41598-018-37186-2
Tryfona T, Bustard M T. J. Biosci. Bioeng., 2008, 105(4): 375-382. doi: 10.1263/jbb.105.375
Julsing M K, Schrewe M, Cornelissen S, et al. Appl. Environ. Microbiol., 2012, 78(16): 5724-5733. doi: 10.1128/AEM.00949-12
Jeon E Y, Song J W, Cha H J, et al. J. Biotechnol., 2018, 281: 161-167. doi: 10.1016/j.jbiotec.2018.07.019
Park H A, Choi K Y. Biochem. Eng. J., 2020, 156: 107524-107532. doi: 10.1016/j.bej.2020.107524
Pagnout C, Sohm B, Razafitianamaharavo A, et al. Sci. Rep., 2019, 9: 1-16. doi: 10.1038/s41598-018-37186-2
Wang Z, Wang J L, Ren G, et al. Marine Drugs, 2015, 13(6): 3325-3339. doi: 10.3390/md13063325
Losey M W, Schmidt M A, Jensen K F. Ind. Eng. Chem. Res., 2001, 40(12): 2555-2562. doi: 10.1021/ie000523f
Miro M, Hansen E H. Anal. Chim. Acta, 2007, 600(1-2): 46-57. doi: 10.1016/j.aca.2007.02.035
Meng S X, Xue L H, Xie C Y, et al. Chem. Eng. J., 2018, 335: 392-400. doi: 10.1016/j.cej.2017.10.085
Xu B B, Zhang Y L, Wei S, et al. ChemCatChem, 2013, 5(8): 2091-2099. doi: 10.1002/cctc.201200863
Znidarsic-Plazl P. J. Flow Chem., 2017, 7(3-4): 111-117.
Hoffmann C, Grundtvig I P R, Thrane J, et al. Chem. Eng. J., 2018, 332: 16-23. doi: 10.1016/j.cej.2017.09.050
Simon D, Obst F, Haefner S, et al. React. Chem. Eng., 2019, 4(1): 67-77. doi: 10.1039/C8RE00180D
Zhu Y, Huang Z, Chen Q, et al. Nat. Commun., 2019, 10: 1-9. doi: 10.1038/s41467-018-07882-8
Piao Y, Han D J, Azad M R, et al. Biosens. Bioelectron., 2015, 65: 220-225. doi: 10.1016/j.bios.2014.10.032
Yang X, Wang Y, Bai R, et al. Green Chem., 2019, 21(9): 2229-2233. doi: 10.1039/C8GC03573C
Sun H, Zheng H, Tang Q, et al. ACS Appl. Mater. Interf., 2019, 11(40): 37313-37321. doi: 10.1021/acsami.9b12479
Stojkovic G, Znidarsic-Plazl P. Acta Chim. Slov., 2010, 57(1): 144-149.
Stojkovic G, Znidarsic-Plazl P. Proc. Biochem., 2012, 47(7): 1102-1107. doi: 10.1016/j.procbio.2012.03.023
Stojkovic G, Krivec M, Vesel A, et al. Appl. Surf. Sci., 2014, 320: 810-817. doi: 10.1016/j.apsusc.2014.09.064
Milozic N, Stojkovic G, Vogel A, et al. New Biotechnol., 2018, 47(25): 18-24.
Liu L S, Kost J, Yan F, et al. Polymers, 2012, 4(2): 997-1011. doi: 10.3390/polym4020997
Zhang J, Yang Z, Li C, et al. Tissue Eng. A, 2013, 19(19-20): 2166-2175. doi: 10.1089/ten.tea.2012.0393
Menegatti T, Znidarsic-Plazl P. Micromachines, 2019, 10(12): 1-12.
Rosche B, Li X Z, Hauer B, et al. Trends Biotechnol., 2009, 27(11): 636-643. doi: 10.1016/j.tibtech.2009.08.001
Halan B, Buehler K, Schmid A. Trends Biotechnol., 2012, 30(9): 453-465. doi: 10.1016/j.tibtech.2012.05.003
Karande R, Debor L, Salamanca D, et al. Biotechnol. Bioeng., 2016, 113(1): 52-61. doi: 10.1002/bit.25696
Willrodt C, Halan B, Karthaus L, et al. Biotechnol. Bioeng., 2017, 114(2): 281-290. doi: 10.1002/bit.26071
Halan B, Karande R, Buehler K, et al. J. Flow Chem., 2016, 6(1): 39-42. doi: 10.1556/1846.2015.00037
Gross R, Lang K, Buehler K, et al. Biotechnol. Bioeng., 2010, 105(4): 705-717.
Zhu C T, Mei Y Y, Zhu L L, et al. Int. J. Mol. Sci., 2018, 19(9): 1-13.
Hoschek A, Heuschkel I, Schmid A, et al. Bioresource Technol., 2019, 282: 171-178. doi: 10.1016/j.biortech.2019.02.093
Roellig R, Plikat C, Ansorge-Schumacher M B. Angew. Chem. Int. Ed., 2019, 58(37): 12960-12963. doi: 10.1002/anie.201907209
Zhao Q, Ansorge-Schumacher M B, Haag R, et al. Bioresource Technol., 2020, 295: 122221. doi: 10.1016/j.biortech.2019.122221

图 2 (a) 微流体反应器系统;(b)微通道中的反应体系
Figure 2 (a) Microfluidic reactor system; (b) Reaction system in microchannel

图 3 (a) L-苹果酸细胞催化的实验装置[51],酵母固定在全氟烷氧基(PFA)微通道的内表面上;(b)实验中的微反应器;(c)固载N-SBM-ATA转化快,固载重组大肠杆菌转化率高,固载ATA几乎无底物转化[53]
Figure 3 (a) Experimental device for L-malic acid cell catalysis [51], and the yeast are immobilized on the inner surface of a perfluoroalkoxy (PFA) microchannel; (b) Experimental microreactor; (c) Immobilization of N-SBM-ATA in microchannels results in fast substrate conversion, immobilization of recombinant E. coli has a high substrate conversion rate, and immobilization ATA has almost no substrate conversion [53]

图 4 (a) 实验中的分段流动生物膜微反应器[59];(b)微通道中分段流入空气和培养基调控生物膜生长[63];(c)微反应器中进行双菌种高细胞密度生物膜培养[64]
Figure 4 (a) Segmented flow biofilm microreactor in experiment [59]; (b) Sectional inflow of air and medium in microchannels to regulate biofilm growth [63]; (c) High cell density biofilm culture of two strains in microreactor [64]

图 5 (a) 手摇形成乳液包封细胞进行生物催化、(b)W/O为2 :8的单隔室乳液和(c)W/O为5 :5的多隔室乳液的激光共聚焦扫描显微镜照片[66]
Figure 5 (a) Hand shake to form emulsion encapsulated cells for biocatalysis; Photo of laser confocal scanning microscope (b) Single-compartment emulsion with W/O of 2 :8, (c) Multi-compartment emulsion with W/O of 5 :5 [66]
表 1 细胞透性化技术强化传质的方法[21~33]
Table 1. Method for enhancing mass transfer by cell permeability technology [21~33]
| 方法 | 减少渗透屏障作用机理 | 特点 | |
| 化学法 | 表面活性剂 | 自发插入细胞膜,降低膜边缘张力,膜上打开小孔并变大,使细胞膜不稳定破裂。 | 较为常用,效果显著,价格较低;易造成细胞破碎,难以重复利用。 |
| 有机溶剂 | 与细胞膜进行静电作用,破坏细胞膜亲疏水平衡,改变膜蛋白结构。 | 常用且效果显著;存在使用安全问题;工艺下游溶剂残留问题。 | |
| 共轭低聚物 | 与细胞表面静电吸引插入磷脂双分子层,改变细胞放电能力,影响细胞Zeta电位。 | 对比表面活性剂和有机溶剂细胞毒性较小;对细胞生长代谢具有抑制作用,重复利用率不高。 | |
| 物理法 | 温度 | 高温:膜蛋白变性形成小孔;冻融:菌体内部产生冰晶破坏细胞膜和细胞壁。 | 高温:方法简便易于放大;不耐高温酶不适用;冻融:对细胞内酶活有利;较为繁琐。 |
| 超声 | 对细胞壁和细胞膜结构进行物理破坏。 | 成本较高;不易放大;会造成酶泄露。 | |
| 电渗透 | 改变膜介电常数使代谢物释放。 | 过程可逆,细胞重复利用率高。 | |
| 分子生物学法 | 分子水平上对膜蛋白成分和表达进行改造。 | 对细胞活性几乎无伤害,细胞重复利用率高。 |
 下载: 导出CSV
下载: 导出CSV

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
