引用本文:
闫琳, 任永硕, 王雪靖, 穆韡, 韩晓军. 凝聚体及其在人造细胞领域中的应用[J]. 化学学报,
2020, 78(11): 1150-1163.
doi:
10.6023/A20060253 Citation:
Yan Lin, Ren Yongshuo, Wang Xuejing, Mu Wei, Han Xiaojun. Coacervate and Its Application in the Field of Artificial Cells[J]. Acta Chimica Sinica,
2020, 78(11): 1150-1163.
doi:
10.6023/A20060253
State Key Laboratory of Urban Water Resource and Environment, School of Chemistry and Chemical Engineering, Harbin Institute of Technology, Harbin 150001
Received Date:
20 June 2020 Available Online:
15 November 2020
Fund Project:
Project supported by the National Natural Science Foundation of China (Nos. 21773050, 21929401) and the Natural Science Foundation of Heilongjiang Province for Distinguished Young Scholars (No. JC2018003)
Abstract:
The origin of life attracts more and more attentions of researchers. Synthetic biologists are devoting to construct a simple and rational system which can exist in primitive earth. Coacervate is a phase separation system which is formed by the interactions of polyelectrolyte. It's a rational protocell model. So far, coacervate has been found to present as membraneless organelles in natural cells. Therefore, the construction of coacervate as artificial organelles is emerging. The formation mechanism, characteristics and categories of coacervate are reviewed in this paper. Additionally, the applications of coacervate as protocell and artificial organelles are summarized. The existing scientific problems and the future development directions are provided at the end of this paper.
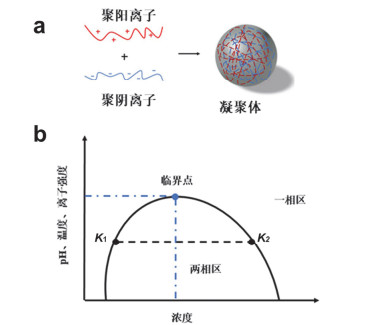
Figure 1.
The formation mechanism and phase diagram of coacervates.
(a) Polycations and polyanions generate coacervates through electrostatic interaction; (b) phase diagram about the effect of concentration, pH, temperature and ionic strength on coacervates formation
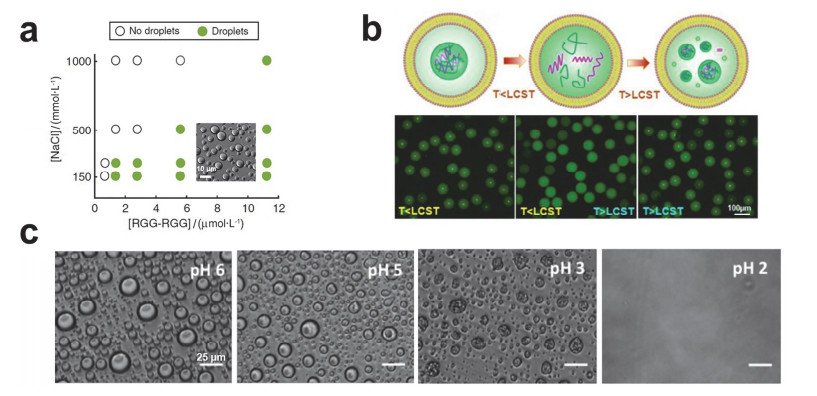
Figure 2.
The effect of ionic strength, temperature and pH on the coacervates.
(a) Phase diagram of (RGG-RGG) protein and salt concentration on coacervates formation[47]. Copyright 2018, Springer Nature. (b) Schematic illustration of the reversible assembly and disassembly of coacervates in liposomes with temperature changeing (top) and the corresponding fluorescence images (bottom) [48]. Copyright 2017, Wiley-VCH. (c) Brightfield micrographs of PGlu/Ply coacervates disassembly by lowering pH [50]. Copyright 2014, American Chemical Society.
Figure 3.
Coacervates with one condensed phase and two coexisting condensed phases.
(a) Schematic illustration of coacervates with one condensed phase[73]; (b) Schematic illustration of coacervates with two coexisting condensed phases[73]; (c) One condensed phase coacervates composed of PDDA and ATP; (d) Two coexisting condensed phases coacervates composed of poly glutamic acid (PGlu)/PAH inner core surrounded by a PGlu/PDDA shell[73]. Copyright 2020, American Chemical Society. https://pubs.acs.org/doi/abs/10.1021/jacs.9b11468
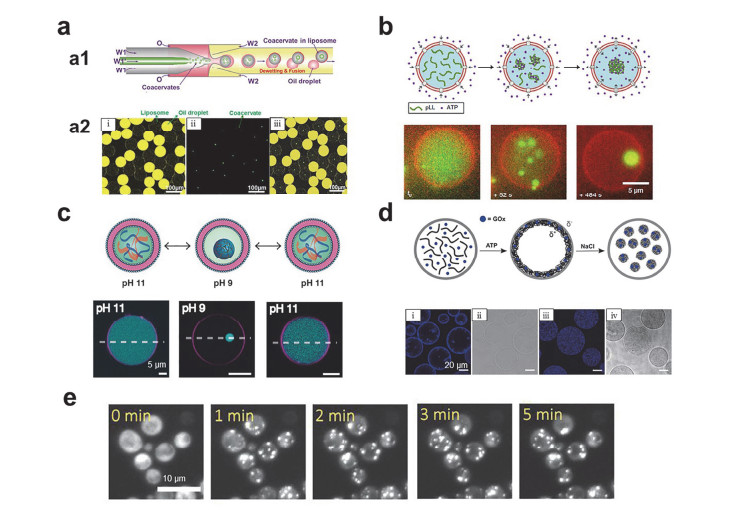
Figure 4.
Application of coacervates as artificial organelles.
(a) Coacervates in vesicles prepared by using microfluidic technology: (a1) Schematic illustration of microfluidic technology, (a2) fluorescence image of (i) oil droplets and vesicles, (ii) coacervates, (iii) the overlap of (i) and (ii) [48]. Copyright 2017, Wiley-VCH. (b) Schematic illustration of the formation of coacervates in vesicles by using α-hemolysin pore (top) and time-lapse fluorescence images (bottom) [117]. Copyright 2019, Springer Nature. (c) Schematic showing that changing the pH to form coacervates reversibly in the vesicles (top) and the corresponding fluorescence images (bottom) [116]. Copyright 2020, Wiley-VCH. (d) Schematic of the formation of coacervates in proteinosomes (top) and fluorescence images: (i, ii) coacervates are adsorbed on the proteinosomes membrane, (iii, iv) coacervates are free in the proteinosomes [112]. Copyright 2019, Wiley-VCH. (e) Optically-regulated protein coacervates formed in a single yeast cell [99]. Copyright 2020, American Chemical Society.
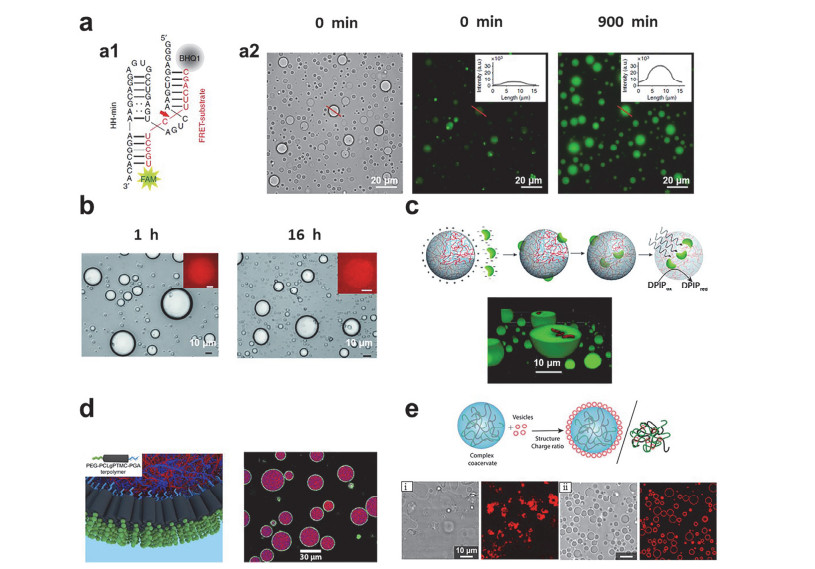
Figure 5.
Individual functions of coacervates artificial cells.
(a) Catalytic cleavage of ribozymes on substrates: (a1) the sequence of hammerhead ribozyme, (a2) Wide-field optical microscopy images of coacervates (left). Fluorescence microscopy images at t=0 min (middle) and t=900 min (right) show an increase in FAM fluorescence (see inset) [127]. Copyright 2018, Springer Nature. (b) Optical microscopy images showing coacervates and recorded 1 h (left) and 16 h (right) after storage in the collection chamber. Insets shows corresponding fluorescence images of mCherry expression in single coacervate[121]. Copyright 2015, Royal Society of Chemistry. (c) Scheme showing the preparation of photosynthetically active membrane-free coacervates containing chloroplasts (top), Confocal fluorescence microscopy images showing coacervates after mixing with a dispersion of intact chloroplasts (bottom) [91]. Copyright 2018, Royal Society of Chemistry. (d) Schematic illustration of copolymer assembly on the surface of the coacervates to form a film (left) and fluorescence images (right): the inset is the schematic of a bespoke terpolymer[68, 69]. Copyright 2017, American Chemical Society. https://pubs.acs.org/doi/full/10.1021/jacs.7b10846. Copyright 2019, American Chemical Society. https://pubs.acs.org/doi/10.1021/acscentsci.9b00345. (e) Schematic illustration of the assembly of phospholipid vesicles on the surface of the coacervates or phospholipid vesicles destroy the coacervates (top) and fluorescence images: (i) the vesicles destroy the coacervates, (ii) the vesicles form a membrane on the surface of the coacervates [130]. Copyright 2019, American Chemical Society.
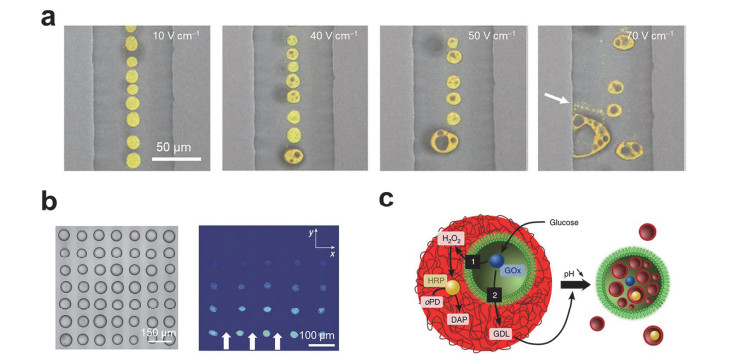
(a) Microscope images of coacervate-based artificial cells under different electric fields[87]. Copyright 2016, Springer Nature. (b) Acoustic patterning of coacervates micro-droplet arrays (left) and the fluorescence microscopy image of transferring of enzyme molecules in the array (right) [137]. Copyright 2018, Springer Nature. (c) Schematic showing synergistic and antagonistic behaviour in synthetic host-guest artificial cells[140]. Copyright 2016, Springer Nature.
Banani, S. F.; Lee, H. O.; Hyman, A. A.; Rosen, M. K. Nat. Rev. Mol. Cell Biol.2017, 18, 285. doi: 10.1038/nrm.2017.7
[14]
Boisvert, F. M.; van Koningsbruggen, S.; Navascues, J.; Lamond, A. I. Nat. Rev. Mol. Cell Biol.2007, 8, 574.
[15]
Gall, J. G. Nat. Rev. Mol. Cell Biol.2003, 4, 975. doi: 10.1038/nrm1262
[16]
Jain, S.; Wheeler, J. R.; Walters, R. W.; Agrawal, A.; Barsic, A.; Parker, R. Cell2016, 164, 487. doi: 10.1016/j.cell.2015.12.038
[17]
Brangwynne, C. P.; Eckmann, C. R.; Courson, D. S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Julicher, F.; Hyman, A. A. Science2009, 324, 1729. doi: 10.1126/science.1172046
Franzmann, T. M.; Jahnel, M.; Pozniakovsky, A.; Mahamid, J.; Holehouse, A. S.; Nuske, E.; Richter, D.; Baumeister, W.; Grill, S. W.; Pappu, R. V.; Hyman, A. A.; Alberti, S. Science2018, 359, eaao5654. doi: 10.1126/science.aao5654
French, J. B.; Jones, S. A.; Deng, H. Y.; Pedley, A. M.; Kim, D.; Chan, C. Y.; Hu, H. B.; Pugh, R. J.; Zhao, H.; Zhang, Y. X.; Huang, T. J.; Fang, Y.; Zhuang, X. W.; Benkovic, S. J. Science2016, 351, 733. doi: 10.1126/science.aac6054
Kim, S.; Huang, J.; Lee, Y.; Dutta, S.; Yoo, H. Y.; Jung, Y. M.; Jho, Y.; Zeng, H.; Hwang, D. S. Proc. Natl. Acad. Sci.2016, 113, E847. doi: 10.1073/pnas.1521521113
[42]
Hoffmann, K. Q.; Perry, S. L.; Leon, L.; Priftis, D.; Tirrell, M.; de Pablo, J. J. Soft Matter2015, 11, 1525. doi: 10.1039/C4SM02336F
[43]
Roy, D.; Brooks, W. L. A.; Sumerlin, B. S. Chem. Soc. Rev.2013, 42, 7214. doi: 10.1039/c3cs35499g
[44]
Blocher, W. C.; Perry, S. L. Wiley Interdiscip. Rev.:Nanomed. Nanobiotechnol. 2017, 9, e1442. doi: 10.1002/wnan.1442
[45]
Priftis, D.; Xia, X.; Margossian, K. O.; Perry, S. L.; Leon, L.; Qin, J.; de Pablo, J. J.; Tirrell, M. Macromolecules2014, 47, 3076. doi: 10.1021/ma500245j
[46]
Jeon, B. J.; Nguyen, D. T.; Abraham, G. R.; Conrad, N.; Fygenson, D. K.; Saleh, O. A. Soft Matter2018, 14, 7009. doi: 10.1039/C8SM01085D
[47]
Schuster, B. S.; Reed, E. H.; Parthasarathy, R.; Jahnke, C. N.; Caldwell, R. M.; Bermudez, J. G.; Ramage, H.; Good, M. C.; Hammer, D. A. Nat. Commun.2018, 9, 12. doi: 10.1038/s41467-017-02416-0
[48]
Deng, N.-N.; Huck W. T. S. Angew. Chem. Int. Ed.2017, 56, 9736. doi: 10.1002/anie.201703145
[49]
Koga, S.; Williams, D. S.; Perriman, A. W.; Mann, S. Nat. Chem.2011, 3, 720. doi: 10.1038/nchem.1110
[50]
Black, K. A.; Priftis, D.; Perry, S. L.; Yip, J.; Byun, W. Y.; Tirrell, M. ACS Macro Lett.2014, 3, 1088. doi: 10.1021/mz500529v
[51]
Aumiller, W. M., Jr.; Keating, C. D. Nat. Chem.2016, 8, 129. doi: 10.1038/nchem.2414
[52]
Spruijt, E.; Sprakel, J.; Stuart, M. A. C.; van der Gucht, J. Soft Matter2010, 6, 172. doi: 10.1039/B911541B
Pathak, J.; Rawat, K. J. Phys. Chem. B2014, 118, 11161. doi: 10.1021/jp5068846
[62]
Perry, S. L.; Leon, L.; Hoffmann, K. Q.; Kade, M. J.; Priftis, D.; Black, K. A.; Wong, D.; Klein, R. A.; Pierce, C. F.; Margossian, K. O.; Whitmer, J. K.; Qin, J.; de Pablo, J. J.; Tirrell, M. Nat. Commun.2015, 6, 8.
[63]
Aumiller, W. M., Jr.; Keating, C. D. Adv. Colloid Interface Sci.2017, 239, 75. doi: 10.1016/j.cis.2016.06.011
[64]
Souza, C. J. F.; da Costa, A. R.; Souza, C. F.; Tosin, F. F. S.; Garcia-Rojas, E. E. Int. J. Biol. Macromol.2018, 107, 1253. doi: 10.1016/j.ijbiomac.2017.09.104
Hwang, D. S.; Zeng, H. B.; Srivastava, A.; Krogstad, D. V.; Tirrell, M.; Israelachvili, J. N.; Waite, J. H. Soft Matter2010, 6, 3232. doi: 10.1039/c002632h
[67]
Zhang, X. M.; Lin, Y. X.; Eschmann, N. A.; Zhou, H. J.; Rauch, J. N.; Hernandez, I.; Guzman, E.; Kosik, K. S.; Han, S. I. PLoS Biol.2017, 15, 28.
[68]
Mason, A. F.; Buddingh, B. C.; Williams, D. S.; van Hest, J. C. M. J. Am. Chem. Soc.2017, 139, 17309. doi: 10.1021/jacs.7b10846
[69]
Mason, A. F.; Yewdall, N. A.; Welzen, P. L. W.; Shao, J.; van Stevendaal, M.; van Hest, J. C. M.; Williams, D. S.; Abdelmohsen, L. ACS Cent. Sci.2019, 5, 1360. doi: 10.1021/acscentsci.9b00345
[70]
Qiao, Y.; Li, M.; Booth, R.; Mann, S. Nat. Chem.2017, 9, 110. doi: 10.1038/nchem.2617
[71]
Dora Tang, T. Y.; Rohaida Che Hak, C.; Thompson, A. J.; Kuimova, M. K.; Williams, D. S.; Perriman, A. W.; Mann, S. Nat. Chem.2014, 6, 527. doi: 10.1038/nchem.1921
Choi, J.-M.; Wang, J.; Holehouse, A. S.; Alberti, S.; Hyman, A. A.; Pappu, R. V. Biophys. J.2018, 114, 561A.
[76]
Wei, M.-T.; Elbaum-Garfinkle, S.; Holehouse, A. S.; Chen, C. C.-H.; Feric, M.; Arnold, C. B.; Priestley, R. D.; Pappu, R. V.; Brangwynne, C. P. Nat. Chem.2017, 9, 1118. doi: 10.1038/nchem.2803
Cohan, M. C.; Posey, A. E.; Mittal, A.; Grigsby, S. J.; Holehouse, A. S.; Buske, P. J.; Levin, P. A.; Pappu, R. V. Mol. Biol. Cell2017, 114, 590A.
[79]
Harmon, T. S.; Holehouse, A. S.; Pappu, R. V. New J. Phys.2018, 20, 045002. doi: 10.1088/1367-2630/aab8d9
[80]
Ruff, K. M.; Pappu, R. V.; Holehouse, A. S. Curr. Opin. Struct. Biol.2019, 56, 1.
[81]
Molliex, A.; Temirov, J.; Lee, J.; Coughlin, M.; Kanagaraj, A. P.; Kim, H. J.; Mittag, T.; Taylor, J. P. Cell2015, 163, 123. doi: 10.1016/j.cell.2015.09.015
[82]
Elbaum-Garfinkle, S.; Kim, Y.; Szczepaniak, K.; Chen, C. C. H.; Eckmann, C. R.; Myong, S.; Brangwynne, C. P. Proc. Natl. Acad. Sci.2015, 112, 7189. doi: 10.1073/pnas.1504822112
[83]
Tan, Y. P.; Hoon, S.; Guerette, P. A.; Wei, W.; Ghadban, A.; Hao, C.; Miserez, A.; Waite, J. H. Nat. Chem. Biol.2015, 11, 488. doi: 10.1038/nchembio.1833
[84]
Madinya, J. J.; Chang, L. W.; Perry, S. L.; Sing, C. E. Mol. Syst. Des. Eng.2020, 5, 632. doi: 10.1039/C9ME00074G
Jing, H.; Bai, Q.; Lin, Y. n.; Chang, H.; Yin, D.; Liang, D. Langmuir2020, 36, 8017. doi: 10.1021/acs.langmuir.0c01864
[87]
Yin, Y. D.; Niu, L.; Zhu, X. C.; Zhao, M. P.; Zhang, Z. X.; Mann, S.; Liang, D. H. Nat. Commun.2016, 7, 7.
[88]
Yin, Y. D.; Chang, H. J.; Jing, H. R.; Zhang, Z. X.; Yan, D. D.; Mann, S.; Liang, D. H. Soft Matter2018, 14, 6514. doi: 10.1039/C8SM01168K
[89]
Fothergill, J.; Li, M.; Davis, S. A.; Cunningham, J. A.; Mann, S. Langmuir2014, 30, 14591. doi: 10.1021/la503746u
[90]
Dompe, M.; Cedano-Serrano, F. J.; Heckert, O.; van den Heuvel, N.; van der Gucht, J.; Tran, Y.; Hourdet, D.; Creton, C.; Kamperman, M. Adv. Mater.2019, 31, e1808179. doi: 10.1002/adma.201808179
[91]
Kumar, B.; Fothergill, J.; Bretherton, J.; Tian, L. F.; Patil, A. J.; Davis, S. A.; Mann, S. Chem. Commun.2018, 54, 3594. doi: 10.1039/C8CC01129J
[92]
Lawrence, M. S.; Phillips, K. J.; Liu, D. R. J. Am. Chem. Soc.2007, 129, 10110. doi: 10.1021/ja071641y
[93]
Comert, F.; Malanowski, A. J.; Azarikia, F.; Dubin, P. L. Soft Matter2016, 12, 4154. doi: 10.1039/C6SM00044D
[94]
Comert, F.; Xu, A. Y.; Madro, S. P.; Liadinskaia, V.; Dubin, P. L. J. Chem. Phys.2018, 149, 163321. doi: 10.1063/1.5029296
[95]
Xu, A. Y.; Melton, L. D.; Ryan, T. M.; Mata, J. P.; Rekas, A.; Williams, M. A. K.; McGillivray, D. J. Food Hydrocoll.2018, 77, 952. doi: 10.1016/j.foodhyd.2017.11.045
[96]
Pandey, P. K.; Kaushik, P.; Rawat, K.; Aswal, V. K.; Bohidar, H. B. Soft Matter2017, 13, 6784. doi: 10.1039/C7SM01222E
Zhang, Y. M.; Rock, C. O. Nat. Rev. Microbiol.2008, 6, 222. doi: 10.1038/nrmicro1839
[108]
Hansen, J. S.; Elbing, K.; Thompson, J. R.; Malmstadt, N.; Lindkvist-Petersson, K. Chem. Commun.2015, 51, 2316. doi: 10.1039/C4CC08838G
[109]
Nordlund, G.; Brzezinski, P.; von Ballmoos, C. Nat. Commun.2014, 5, 8.
[110]
Lee, K. Y.; Park, S. J.; Lee, K. A.; Kim, S. H.; Kim, H.; Meroz, Y.; Mahadevan, L.; Jung, K. H.; Ahn, T. K.; Parker, K. K.; Shin, K. Nat. Biotechnol.2018, 36, 530. doi: 10.1038/nbt.4140
[111]
Sokolova, E.; Spruijt, E.; Hansen, M. M. K.; Dubuc, E.; Groen, J.; Chokkalingam, V.; Piruska, A.; Heus, H. A.; Huck, W. T. S. Proc. Natl. Acad. Sci.2013, 110, 11692. doi: 10.1073/pnas.1222321110
[112]
Booth, R.; Qiao, Y.; Li, M.; Mann, S. Angew. Chem. Int. Ed.2019, 3, 1.
[113]
Last, M. G. F.; Deshpande, S.; Dekker, C. ACS Nano2020, 14, 4487. doi: 10.1021/acsnano.9b10167
[114]
Sato, Y.; Takinoue, M. Micromachines2019, 10, 216. doi: 10.3390/mi10040216
[115]
Linsenmeier, M.; Kopp, M. R. G.; Grigolato, F.; Liu, D.; Zuercher, D.; Hondele, M.; Weis, K.; Palmiero, U. C.; Arosio, P. Angew. Chem. Int. Ed.2019, 58, 14489. doi: 10.1002/anie.201907278
[116]
Love, C.; Steinkuhler, J.; Gonzales, D. T.; Yandrapalli, N.; Robinson, T.; Dimova, R.; Tang, T. D. Angew. Chem. Int. Ed.2020, 59, 5950. doi: 10.1002/anie.201914893
[117]
Deshpande, S.; Brandenburg, F.; Lau, A.; Last, M. G. F.; Spoelstra, W. K.; Reese, L.; Wunnava, S.; Dogterom, M.; Dekker, C. Nat. Commun.2019, 10, 1800. doi: 10.1038/s41467-019-09855-x
[118]
Shin, Y.; Berry, J.; Pannucci, N.; Haataja, M. P.; Toettcher, J. E.; Brangwynne, C. P. Cell2017, 168, 159. doi: 10.1016/j.cell.2016.11.054
[119]
Yewdall, N. A.; Mason, A. F.; van Hest, J. C. M. Interface Focus2018, 8, 15.
[120]
Zwicker, D.; Seyboldt, R.; Weber, C. A.; Hyman, A. A.; Jülicher, F. Nat. Phys.2016, 13, 408.
[121]
Tang, T. Y. D.; van Swaay, D.; deMello, A.; Anderson, J. L. R.; Mann, S. Chem. Commun.2015, 51, 11429. doi: 10.1039/C5CC04220H
[122]
Deng, N.-N.; Vibhute, M. A.; Zheng, L.; Zhao, H.; Yelleswarapu, M.; Huck, W. T. S. J. Am. Chem. Soc.2018, 140, 7399. doi: 10.1021/jacs.8b03123
[123]
Krishna Kumar, R.; Yu, X.; Patil, A. J.; Li, M.; Mann, S. Angew. Chem. Int. Ed.2011, 50, 9343. doi: 10.1002/anie.201102628
[124]
Dzieciol, A. J.; Mann, S. Chem. Soc. Rev.2012, 41, 79. doi: 10.1039/C1CS15211D
[125]
Chatterjee, S.; Yadav, S. Life2019, 9, 1.
[126]
Strulson, C. A.; Molden, R. C.; Keating, C. D.; Bevilacqua, P. C. Nat. Chem.2012, 4, 941. doi: 10.1038/nchem.1466
[127]
Drobot, B.; Iglesias-Artola, J. M.; Le Vay, K.; Mayr, V.; Kar, M.; Kreysing, M.; Mutschler, H.; Tang, T. D. Nat. Commun.2018, 9, 3643. doi: 10.1038/s41467-018-06072-w
[128]
Poudyal, R. R.; Guth-Metzler, R. M.; Veenis, A. J.; Frankel, E. A.; Keating, C. D.; Bevilacqua, P. C. Nat. Commun.2019, 10, 490. doi: 10.1038/s41467-019-08353-4
[129]
Tang, T. Y. D.; Antognozzi, M.; Vicary, J. A.; Perriman, A. W.; Mann, S. Soft Matter2013, 9, 7647. doi: 10.1039/c3sm50726b
Martin, N.; Douliez, J. P. Angew. Chem. Int. Ed.2017, 56, 13689. doi: 10.1002/anie.201707139
[132]
Williams, D. S.; Patil, A. J.; Mann, S. Small2014, 10, 1830. doi: 10.1002/smll.201303654
[133]
Byun, C. K.; Hwang, H.; Choi, W. S.; Yaguchi, T.; Park, J.; Kim, D.; Mitchell, R. J.; Kim, T.; Cho, Y. K.; Takayama, S. J. Am. Chem. Soc.2013, 135, 2242. doi: 10.1021/ja3094923
Wang, X. J.; Tian, L. F.; Du, H.; Li, M.; Mu, W.; Drinkwater, B. W.; Han, X. J.; Mann, S. Chem. Sci.2019, 10, 9446. doi: 10.1039/C9SC04522H
[136]
Magdalena Estirado, E.; Mason, A. F.; Alemán García, M. Á.; van Hest, J. C. M.; Brunsveld, L. J. Am. Chem. Soc.2020, 142, 9106. doi: 10.1021/jacs.0c01732
[137]
Tian, L.; Martin, N.; Bassindale, P. G.; Patil, A. J.; Li, M.; Barnes, A.; Drinkwater, B. W.; Mann, S. Nat. Commun.2016, 7, 13068. doi: 10.1038/ncomms13068
[138]
Tian, L. F.; Li, M.; Liu, J. T.; Patil, A. J.; Drinkwater, B. W.; Mann, S. ACS Cent. Sci.2018, 4, 1551. doi: 10.1021/acscentsci.8b00555
[139]
Tian, L. F.; Li, M.; Patil, A. J.; Drinkwater, B. W.; Mann, S. Nat. Commun.2019, 10, 13. doi: 10.1038/s41467-018-07689-7
[140]
Martin, N.; Douliez, J. P.; Qiao, Y.; Booth, R.; Li, M.; Mann, S. Nat. Commun.2018, 9, 3652. doi: 10.1038/s41467-018-06087-3
[141]
Qiao, Y.; Li, M.; Qiu, D.; Mann, S. Angew. Chem. Int. Ed.2019, 58, 17758. doi: 10.1002/anie.201909313
[142]
Zhang, Y. W.; Liu, S. Y.; Yao, Y.; Chen, Y. F.; Zhou, S. H.; Yang, X. H.; Wang, K.; Liu, J. B. Small2020, 16, 2002073. doi: 10.1002/smll.202002073
te Brinke, E.; Groen, J.; Herrmann, A.; Heus, H. A.; Rivas, G.; Spruijt, E.; Huck, W. T. S. Nat. Nanotechnol.2018, 13, 849. doi: 10.1038/s41565-018-0192-1
[147]
Zwicker, D.; Seyboldt, R.; Weber, C. A.; Hyman, A. A.; Julicher, F. Nat. Phys.2017, 13, 408. doi: 10.1038/nphys3984
[148]
Spoelstra, W. K.; van der Sluis, E. O.; Dogterom, M.; Reese, L. Langmuir2020, 36, 1956. doi: 10.1021/acs.langmuir.9b02719
图 1
凝聚体的形成机理及相图.
Figure 1
The formation mechanism and phase diagram of coacervates.
(a) Polycations and polyanions generate coacervates through electrostatic interaction; (b) phase diagram about the effect of concentration, pH, temperature and ionic strength on coacervates formation
(a) Phase diagram of (RGG-RGG) protein and salt concentration on coacervates formation[47]. Copyright 2018, Springer Nature. (b) Schematic illustration of the reversible assembly and disassembly of coacervates in liposomes with temperature changeing (top) and the corresponding fluorescence images (bottom) [48]. Copyright 2017, Wiley-VCH. (c) Brightfield micrographs of PGlu/Ply coacervates disassembly by lowering pH [50]. Copyright 2014, American Chemical Society.
(a) Schematic illustration of coacervates with one condensed phase[73]; (b) Schematic illustration of coacervates with two coexisting condensed phases[73]; (c) One condensed phase coacervates composed of PDDA and ATP; (d) Two coexisting condensed phases coacervates composed of poly glutamic acid (PGlu)/PAH inner core surrounded by a PGlu/PDDA shell[73]. Copyright 2020, American Chemical Society. https://pubs.acs.org/doi/abs/10.1021/jacs.9b11468
(a) Coacervates in vesicles prepared by using microfluidic technology: (a1) Schematic illustration of microfluidic technology, (a2) fluorescence image of (i) oil droplets and vesicles, (ii) coacervates, (iii) the overlap of (i) and (ii) [48]. Copyright 2017, Wiley-VCH. (b) Schematic illustration of the formation of coacervates in vesicles by using α-hemolysin pore (top) and time-lapse fluorescence images (bottom) [117]. Copyright 2019, Springer Nature. (c) Schematic showing that changing the pH to form coacervates reversibly in the vesicles (top) and the corresponding fluorescence images (bottom) [116]. Copyright 2020, Wiley-VCH. (d) Schematic of the formation of coacervates in proteinosomes (top) and fluorescence images: (i, ii) coacervates are adsorbed on the proteinosomes membrane, (iii, iv) coacervates are free in the proteinosomes [112]. Copyright 2019, Wiley-VCH. (e) Optically-regulated protein coacervates formed in a single yeast cell [99]. Copyright 2020, American Chemical Society.
(a) Catalytic cleavage of ribozymes on substrates: (a1) the sequence of hammerhead ribozyme, (a2) Wide-field optical microscopy images of coacervates (left). Fluorescence microscopy images at t=0 min (middle) and t=900 min (right) show an increase in FAM fluorescence (see inset) [127]. Copyright 2018, Springer Nature. (b) Optical microscopy images showing coacervates and recorded 1 h (left) and 16 h (right) after storage in the collection chamber. Insets shows corresponding fluorescence images of mCherry expression in single coacervate[121]. Copyright 2015, Royal Society of Chemistry. (c) Scheme showing the preparation of photosynthetically active membrane-free coacervates containing chloroplasts (top), Confocal fluorescence microscopy images showing coacervates after mixing with a dispersion of intact chloroplasts (bottom) [91]. Copyright 2018, Royal Society of Chemistry. (d) Schematic illustration of copolymer assembly on the surface of the coacervates to form a film (left) and fluorescence images (right): the inset is the schematic of a bespoke terpolymer[68, 69]. Copyright 2017, American Chemical Society. https://pubs.acs.org/doi/full/10.1021/jacs.7b10846. Copyright 2019, American Chemical Society. https://pubs.acs.org/doi/10.1021/acscentsci.9b00345. (e) Schematic illustration of the assembly of phospholipid vesicles on the surface of the coacervates or phospholipid vesicles destroy the coacervates (top) and fluorescence images: (i) the vesicles destroy the coacervates, (ii) the vesicles form a membrane on the surface of the coacervates [130]. Copyright 2019, American Chemical Society.
(a) Microscope images of coacervate-based artificial cells under different electric fields[87]. Copyright 2016, Springer Nature. (b) Acoustic patterning of coacervates micro-droplet arrays (left) and the fluorescence microscopy image of transferring of enzyme molecules in the array (right) [137]. Copyright 2018, Springer Nature. (c) Schematic showing synergistic and antagonistic behaviour in synthetic host-guest artificial cells[140]. Copyright 2016, Springer Nature.

 下载:
下载:


 下载:
下载:







