-
[1]
Petersen P E. Oral Cancer Prevention and Control-the Approach of the World Health Organization[J]. Oral Oncol,
2009, 45(4/5):
454-460.
-
[2]
Blagih J, Coulombe F, Vincent E E. The Energy Sensor AMPK Regulates T Cell Metabolic Adaptation and Effector Responses in Vivo[J]. Immunity,
2015, 42(1):
41-54.
doi: 10.1016/j.immuni.2014.12.030
-
[3]
张继东, 李家源, 金华峰. 基于荧光效应的小分子抗癌药物释放体系研究进展[J]. 应用化学,
2019,36,(7): 733-748.
ZHANG Jidong, LI Jiayuan, JIN Huafeng. Research Progress on Small Molecule Anticancer Drug Release System Based on Fluorescence Effect[J]. Chinese J Appl Chem,
2019, 36(7):
733-748.
-
[4]
王玉鹏, 周东方, 程延祥. 血红蛋白/光敏剂复合药物体系用于光动力治疗[J]. 应用化学,
2018,35,(12): 1442-1448.
WANG Yupeng, ZHOU Dongfang, CHENG Yanxiang. Hemoglobin/Photosensitizer Compound Drug System for Photodynamic Therapy[J]. Chinese J Appl Chem,
2018, 35(12):
1442-1448.
-
[5]
Dougan M, Dougan S K. Targeting Immunotherapy to the Tumor Microenvironment[J]. J Cell Biochem,
2017, 118:
3049-3054.
doi: 10.1002/jcb.26005
-
[6]
Palucka K, Banchereau J. Cancer Immunotherapy via Dendritic Cells[J]. Nat Rev Cancer,
2012, 12(4):
265-277.
doi: 10.1038/nrc3258
-
[7]
McNutt M. Cancer Immunotherapy[J]. Science,
2013, 342(6165):
1417.
doi: 10.1126/science.1249481
-
[8]
Schumacher T N, Schreiber R D. Neoantigens in Cancer Immunotherapy[J]. Science,
2015, 348(6230):
69-74.
doi: 10.1126/science.aaa4971
-
[9]
Sharma P, Allison J P. The Future of Immune Checkpoint Therapy[J]. Science,
2015, 348(6230):
56-61.
doi: 10.1126/science.aaa8172
-
[10]
Wu X, Giobbie-Hurder A, Liao X. Angiopoietin-2 as a Biomarker and Target for Immune Checkpoint Therapy[J]. Cancer Immunol Res,
2017, 5(1):
17-28.
-
[11]
Yarchoan M, Hopkins A, Jaffee E M. Tumor Mutational Burden and Response Rate to PD-1 Inhibition[J]. New Engl J Med,
2017, 377(25):
2500-2501.
doi: 10.1056/NEJMc1713444
-
[12]
Santarpia M, Karachaliou N. Tumor Immune Microenvironment Characterization and Response to Anti-PD-1 Therapy[J]. Cancer Biol Med,
2015, 12(2):
74-78.
-
[13]
Netea-Maier R T, Smit J W A, Netea M G. Metabolic Changes in Tumor Cells and Tumor-Associated Macrophages:A Mutual Relationship[J]. Cancer Lett,
2018, 413:
102-109.
doi: 10.1016/j.canlet.2017.10.037
-
[14]
Yu L, Chen X, Wang L. The Sweet Trap in Tumors:Aerobic Glycolysis and Potential Targets for Therapy[J]. Oncotarget,
2016, 7(25):
38908-38926.
doi: 10.18632/oncotarget.7676
-
[15]
Warburg O, Wind F, Negelein E. The Metabolism of Tumors in the Body[J]. J Gen Physiol,
1927, 8:
519-530.
doi: 10.1085/jgp.8.6.519
-
[16]
Gatenby R A, Gillies R J. Why do Cancers Have High Aerobic Glycolysis?[J]. Nat Rev Cancer,
2004, 4(11):
891-899.
doi: 10.1038/nrc1478
-
[17]
Porporato P E, Payen V L, Baselet B. Metabolic Changes Associated with Tumor Metastasis, Part 2:Mitochondria, Lipid and Amino Acid Metabolism[J]. Cell Mol Life Sci,
2016, 73(7):
1349-1363.
doi: 10.1007/s00018-015-2100-2
-
[18]
Currie E, Schulze A, Zechner R. Cellular Fatty Acid Metabolism and Cancer[J]. Cell Metab,
2013, 18(2):
153-161.
-
[19]
Ho P C, Liu P S. Metabolic Communication in Tumors:A New Layer of Immunoregulation for Immune Evasion[J]. J Immunother Cancer,
2016, 4(1):
4.
-
[20]
Lunt S Y, Vander Heiden M G. Aerobic Glycolysis:Meeting the Metabolic Requirements of Cell Proliferation[J]. Annu Rev Cell Dev Bi,
2011, 27:
441-464.
doi: 10.1146/annurev-cellbio-092910-154237
-
[21]
Li Z, Zhang H. Reprogramming of Glucose, Fatty Acid and Amino Acid Metabolism for Cancer Progression[J]. Cell Mol Life Sci,
2016, 73(2):
377-392.
doi: 10.1007/s00018-015-2070-4
-
[22]
Ho P C, Bihuniak J D, Macintyre A N. Phosphoenolpyruvate Is a Metabolic Checkpoint of Anti-tumor T Cell Responses[J]. Cell,
2015, 162(6):
1217-1228.
doi: 10.1016/j.cell.2015.08.012
-
[23]
Yang B, Ding L, Chen Y. Augmenting Tumor-Starvation Therapy by Cancer Cell Autophagy Inhibition[J]. Adv Sci,
2020, 7(6):
1902847.
doi: 10.1002/advs.201902847
-
[24]
Xie W, Deng W W, Zan M. Cancer Cell Membrane Camouflaged Nanoparticles to Realize Starvation Therapy Together with Checkpoint Blockades for Enhancing Cancer Therapy[J]. ACS Nano,
2019, 13(3):
2849-2857.
-
[25]
Gao F, Cheng Q, Liu M D. Local T Regulatory Cells Depletion by an Integrated Nanodrug System for Efficient Chem-Immunotherapy of Tumor[J]. Sci China Chem,
2019, 62(9):
1230-1244.
doi: 10.1007/s11426-019-9507-x
-
[26]
Jiang B. Aerobic Glycolysis and High Level of Lactate in Cancer Metabolism and Microenvironment[J]. Gene Dis,
2017, 4(1):
25-27.
doi: 10.1016/j.gendis.2017.02.003
-
[27]
Gao F, Tang Y, Liu W L. Intra/Extracellular Lactic Acid Exhaustion for Synergistic Metabolic Therapy and Immunotherapy of Tumors[J]. Adv Mater,
2019, 31(51):
1904639.
doi: 10.1002/adma.201904639
-
[28]
Sukumar M, Liu J, Ji Y. Inhibiting Glycolytic Metabolism Enhances CD8+ T Cell Memory and Antitumor Function[J]. J Clin Invest,
2013, 123(10):
4479-4488.
doi: 10.1172/JCI69589
-
[29]
Labow B I, Souba W W. Glutamine[J]. World J Surg,
2000, 24(12):
1503-1513.
doi: 10.1007/s002680010269
-
[30]
Shanware N P, Mullen A R, DeBerardinis R J. Glutamine:Pleiotropic Roles in Tumor Growth and Stress Resistance[J]. J Mol Med,
2011, 89(3):
229-236.
-
[31]
Klysz D, Tai X G, Robert P A. Glutamine-Dependent Alpha-Ketoglutarate Production Regulates the Balance Between T Helper 1 Cell and Regulatory T Cell Generation[J]. Sci Signal,
2015, 8(396):
ra97.
doi: 10.1126/scisignal.aab2610
-
[32]
Nabe S, Yamada T, Suzuki J. Reinforce the Antitumor Activity of CD8+ T Cells via Glutamine Restriction[J]. Cancer Sci,
2018, 109(12):
3737-3750.
doi: 10.1111/cas.13827
-
[33]
Leone R D, Zhao L, Englert J M. Glutamine Blockade Induces Divergent Metabolic Programs to Overcome Tumor Immune Evasion[J]. Science,
2019, 366(6468):
1013-1021.
doi: 10.1126/science.aav2588
-
[34]
Sukumar M, Liu J, Ji Y. Inhibiting Glycolytic Metabolism Enhances CD8+ T Cell Memory and Antitumor Function[J]. J Clin Invest,
2013, 123(10):
4479-4488.
doi: 10.1172/JCI69589
-
[35]
Greten T F, Manns M P, Korangy F. Myeloid Derived Suppressor Cells in Human Diseases[J]. Int Immunopharmacol,
2011, 11(7):
802-807.
doi: 10.1016/j.intimp.2011.01.003
-
[36]
Li X Y, Wenes M, Romero P. Navigating Metabolic Pathways to Enhance Antitumor Immunity and Immunotherapy[J]. Nat Rev Clin Oncol,
2019, 16(7):
425-441.
doi: 10.1038/s41571-019-0203-7
-
[37]
Norian L A, Rodriguez P C, O'Mara L A. Tumor-Infiltrating Regulatory Dendritic Cells Inhibit CD8+ T Cell Function via L-Arginine Metabolism[J]. Cancer Res,
2009, 69(7):
3086-3094.
doi: 10.1158/0008-5472.CAN-08-2826
-
[38]
Brin E, Wu K, Lu H T. PEGylated Arginine Deiminase can Modulate Tumor Immune Microenvironment by Affecting Immune Checkpoint Expression, Decreasing Regulatory T Cell Accumulation and Inducing Tumor T Cell Infiltration[J]. Oncotarget,
2017, 8:
58948-58963.
doi: 10.18632/oncotarget.19564
-
[39]
Geiger R, Rieckmann J C, Wolf T. L-Arginine Modulates T Cell Metabolism and Enhances Survival and Anti-Tumor Activity[J]. Cell,
2016, 167:
829-842.
doi: 10.1016/j.cell.2016.09.031
-
[40]
Ron-Harel N, Santos D, Ghergurovich J M. Mitochondrial Biogenesis and Proteome Remodeling Promote One-Carbon Metabolism for T Cell Activation[J]. Cell Metab,
2016, 24(1):
104-117.
-
[41]
Cheng C M, Geng F, Cheng X. Lipid Metabolism Reprogramming and Its Potential Targets in Cancer[J]. Cancer Commun,
2018, 38(1):
27.
-
[42]
Monaco M E. Fatty Acid Metabolism in Breast Cancer Subtypes[J]. Oncotarget,
2017, 8(17):
29487-29500.
doi: 10.18632/oncotarget.15494
-
[43]
Liu Y. Fatty Acid Oxidation is a Dominant Bioenergetic Pathway in Prostate Cancer[J]. Prostate Cancer P D,
2006, 9(3):
230-234.
-
[44]
Clement E, Lazar I, Attan C. Adipocyte Extracellular Vesicles Carry Enzymes and Fatty Acids that Stimulate Mitochondrial Metabolism and Remodeling in Tumor Cells[J]. EMBO J,
2020, 39(3):
e102525.
-
[45]
Pearce E L, Walsh M C, Cejas P J. Enhancing CD8 T-Cell Memory by Modulating Fatty Acid Metabolism[J]. Nature,
2009, 460(7251):
103-107.
doi: 10.1038/nature08097
-
[46]
Pacella I, Procaccini C, Focaccetti C. Fatty Acid Metabolism Complements Glycolysis in the Selective Regulatory T Cell Expansion During Tumor Growth[J]. P Natl Acad Sci USA,
2018, 115(28):
E6546-E6555.
doi: 10.1073/pnas.1720113115
-
[47]
Zhang Y, Kurupati R, Liu L. Enhancing CD8+ T Cell Fatty Acid Catabolism within a Metabolically Challenging Tumor Microenvironment Increases the Efficacy of Melanoma Immunotherapy[J]. Cancer Cell,
2017, 32:
377-391.
doi: 10.1016/j.ccell.2017.08.004
-
[48]
Zhang C, Yue C, Herrmann A. STAT3 Activation-Induced Fatty Acid Oxidation in CD8+ T Effector Cells is Critical for Obesity-Promoted Breast Tumor Growth[J]. Cell Metab,
2020, 31(1):
148-161.
-
[49]
Xia J L, Tian H Y, Chen J. Polyglutamic Acid Based Polyanionic Shielding System for Polycationic Gene Carriers[J]. Chinese J Polym Sci,
2016, 34(3):
316-323.
-
[50]
林琳, 郭兆培, 陈杰. 聚苯丙氨酸修饰低分子量聚乙烯亚胺制备高效基因载体[J]. 高分子学报,
2017,2,(2): 321-328.
LIN Lin, GUO Zhaopei, CHEN Jie. Synthesis and Characterization of Polyphenylalanine Grafted Low Molecular Weight PEI as Efficient Gene Carriers[J]. Acta Polym Sin,
2017, 2(2):
321-328.
-
[51]
Xu C N, Tian H Y, Wang Y B. Anti-tumor Effects of Combined Doxorubicin and siRNA for Pulmonary Delivery[J]. Chinese Chem Lett,
2017, 28(4):
807-812.
doi: 10.1016/j.cclet.2016.12.013
-
[52]
Zhao H, Xu J, Li Y. Nanoscale Coordination Polymer Based Nanovaccine for Tumor Immunotherapy[J]. ACS Nano,
2019, 13(11):
13127-13135.
doi: 10.1021/acsnano.9b05974

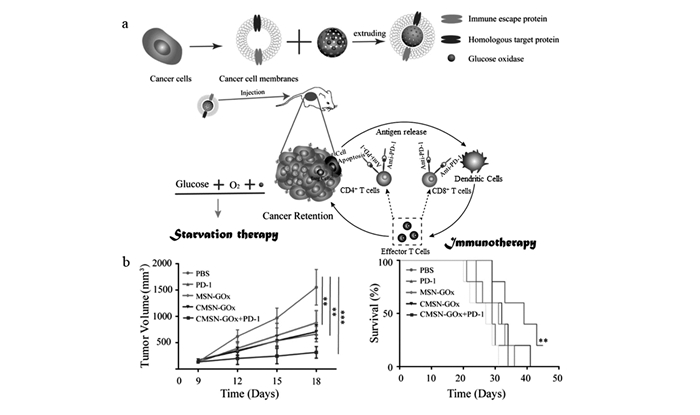
 下载:
下载:
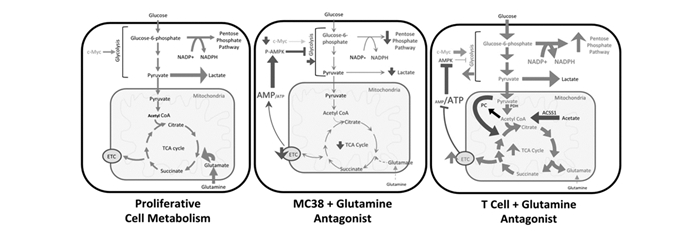
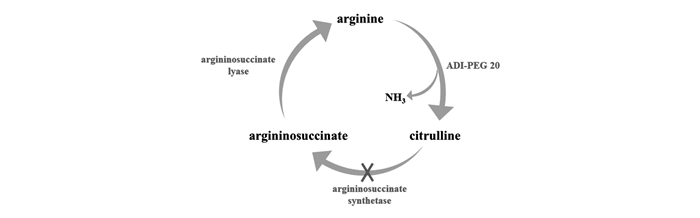
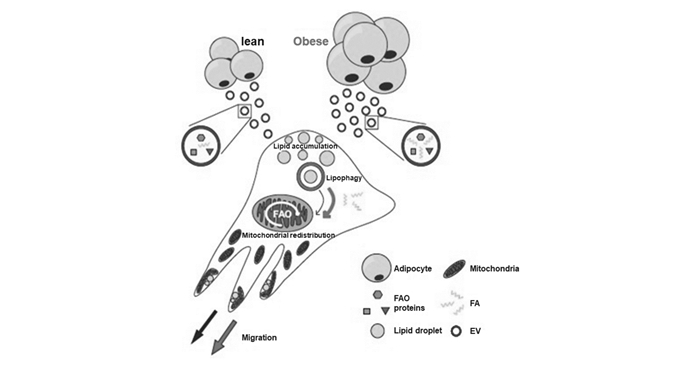

 下载:
下载:





