Citation:
Zhilong Xie, Guohui Zhang, Ya Meng, Yefei Tong, Jian Deng, Honghui Li, Qingqing Ma, Shisong Han, Wenjun Ni. A natural nano-platform: Advances in drug delivery system with recombinant high-density lipoprotein[J]. Chinese Chemical Letters,
2024, 35(11): 109584.
doi:
10.1016/j.cclet.2024.109584
A natural nano-platform: Advances in drug delivery system with recombinant high-density lipoprotein
English
A natural nano-platform: Advances in drug delivery system with recombinant high-density lipoprotein
Zhuhai Clinical Medical College of Jinan University (The Department of Urology, Zhuhai People's Hospital), Zhuhai 519050, China
b.
Guangdong Provincial Key Laboratory of Tumor Interventional Diagnosis and Treatment, Zhuhai Institute of Translational Medicine, Zhuhai People's Hospital (Zhuhai Clinical Medical College of Jinan University), Zhuhai 519050, China
c.
The Second Clinical Medical College, Guangdong Medical University, Dongguan 523808, China
d.
Guizhou Aerospace Hospital, Guizhou 563099, China
* Corresponding authors. E-mail addresses: hanshs@qq.com (S. Han)
niwj5@jnu.edu.cn (W. Ni). 1 These authors contributed equally to this work.
Received Date:
02 December 2023 Accepted Date:
22 January 2024 Revised Date:
19 January 2024 Available Online:
15 November 2024
Abstract:
Developing natural nano-platforms with high biocompatibility and natural targeting ability represents great significance for drug delivery. High-density lipoprotein (HDL), a natural lipid-protein complex, plays important roles in physiological activities, particularly in reverse cholesterol transport (RCT) and be closely associated with atherosclerotic cardiovascular diseases. Recent studies have demonstrated that HDLs have the potential to serve as ideal drug carriers. Recombinant HDLs (rHDLs) have been used to encapsulate substances such as small interfering RNA (siRNA), drugs, and contrast agents, fully utilizing the biocompatibility and targeting ability of rHDL in the body and providing new strategies for drug delivery and disease treatment. In this review, we discussed in detail the basic principles of HDL as a drug delivery system, the mechanisms of targeted drug delivery, and several methods for preparing HDL nanoparticles. Afterward, we comprehensively reviewed the applications of HDL as a drug carrier in cardiovascular diseases, cancer treatment (such as glioblastoma, breast cancer, hepatocellular carcinoma and urologic cancers) and some other fields. Finally, we reviewed the therapeutic effects and safety of HDL nanoparticles in clinical studies. Through a review and summary of these research advances, we aim to fully understand the potential of HDL as a drug carrier in clinical applications, providing valuable references and guidance for future research and expedites the translational application of HDL as drug carriers.
High-density lipoprotein (HDL) is a complex lipid-protein composite that plays a crucial biological role in the body. In recent decades, HDL has been referred to as the "good cholesterol" due to its pivotal function in reverse cholesterol transport, which is important in combating atherosclerosis [1]. Since Wilson et al. reported a close association between plasma HDL levels and cardiovascular disease in 1980 [2], research on the relationship between plasma lipoproteins and cardiovascular disease has continued. The effects of HDL levels on the occurrence of cardiovascular disease have been extensively studied and elucidated. As of 2023, the utilization of nanoparticle-gated electrokinetic membrane sensors (NGEMS) has allowed the quantitative assessment of paraoxonase 1 (PON 1)-HDL in plasma, enabling simple and rapid evaluation of the risk of cardiovascular disease [3]. As research has advanced, other functions of HDL have gradually been discovered. In addition to its role in cholesterol metabolism and transport, HDL possesses various biological activities, including antioxidant, anti-inflammatory, and antiplatelet aggregation properties [4-7]. HDL is a natural nanomaterial in the human body with a diameter of approximately 8–12 nm. It is non-immunogenic, meaning that the body's reticuloendothelial system (RES) cannot easily recognize or clear it. Moreover, HDL has a circulatory half-life of 12–24 h and targets different receptor cells in the body [8].
In recent years, researchers have focused on exploring nanoparticles capable of integrating drug delivery and targeted release, leading to the development of various nanomaterials. Liposomes are one such type, consisting of a bilayer membrane composed of phospholipids and cholesterol molecules. Its biological properties are roughly analogous to those of HDL. The use of liposomes as drug carriers has successfully achieved transmembrane transport [9]. Simultaneously, researchers have found that the integration of dual-doped carbon quantum dots (CQDs) with liposomes can yield effects conducive to fluorescence visualization and therapy. This has propelled the application of liposomes [10]. Despite the drug-carrying and release properties of liposomes, their passive targeting characteristics limit the precision of drug delivery. In addition, micelles also exhibit excellent performance as drug carriers. For instance, various shapes of micelles can be employed to enhance intestinal mucus diffusion and drug accumulation [11].
However, an increasing body of research has shown that HDL not only plays an indispensable role in the occurrence and development of cardiovascular diseases, but also holds potential as a drug carrier, offering new strategies for drug delivery and disease treatment. Several laboratories have successfully utilized HDL as a carrier to transport specific drugs to target tissues, facilitating drug accumulation and enhancing disease treatment efficiency. For instance, Ouyang et al. successfully constructed a biomimetic recombinant high-density lipoprotein (rHDL/CD-PEI/P53 complex) drug delivery system targeting scavenger receptor class B type I SR-BI, which demonstrated excellent therapeutic effects in bladder cancer [12]. Subsequently, researchers encapsulated the anticancer drug valrubicin in superparamagnetic iron oxide nanoparticles (SPIONs) containing recombinant HDL (rHDL). Through the combination of iron nanoparticles and HDL, targeted drug delivery to the prostate was achieved, leading to the inhibition of prostate cancer cell growth and spread [13]. Traditionally, drug delivery has often relied on the synthesis of other inorganic nanoparticles or carriers. However, these carriers have some limitations in terms of stability, biocompatibility, and targeting ability in the body [14]. Although researchers have now developed numerous stimuli-responsive nano-drug delivery systems [15]. In contrast, HDL, a naturally occurring lipid-protein complex, has unique advantages. Its stability and biocompatibility in the body have been widely recognized, and the binding of HDL to specific receptors enables the precise targeting of specific tissues or cells [16]. The potential of HDL as a drug carrier extends beyond drug delivery and includes its application in therapies. Because of its crucial role in inflammatory responses and immune regulation, HDL can be used as a drug carrier for the treatment of inflammatory and immune-related diseases. Moreover, HDL participates in the treatment of cardiovascular diseases by modulating cholesterol metabolism and clearing oxidized low-density lipoprotein (LDL) [17].
In this review, we introduce the structure, synthesis, and biological characteristics of HDL and delve into its relationship with cholesterol metabolism and cardiovascular diseases. Subsequently, we explain in detail the fundamental principles of HDL as a drug delivery system and the mechanisms of targeted drug delivery. We explore the differences between HDL and other nanoparticles and review the preparation methods for these nanoparticles. The crucial aspect is that we comprehensively summarize the applications of HDL as a drug carrier in cardiovascular diseases, cancer treatment, and other fields. Through exploration of these research advancements, we can gain a better understanding of the potential clinical applications of HDL as a drug carrier. Finally, we reviewed the therapeutic effects and safety of HDL nanoparticles in clinical studies. This study provides valuable references and guidance for future research and expedites the translational application of HDL as drug carriers. However, we must also recognize the research on HDL as a drug carrier faces challenges and limitations. These include improvements in nanoparticle preparation techniques, optimization of drug packaging and release systems, and in-depth research into drug delivery, efficacy, and safety. We anticipate that further advances and applications of HDL as a drug carrier will be achieved through these efforts.
In conclusion, this review aims to comprehensively present the research progress of high-density lipoprotein as drug carriers. By examining the biological characteristics, principles of drug delivery, and application examples of HDL, we sought to gain a comprehensive understanding of its significance and potential as a drug carrier. We firmly believe that research on HDL as a drug carrier will bring new breakthroughs in the drug delivery and therapeutic fields, providing more precise and effective strategies for disease treatment.
2.
Biological properties of HDLs
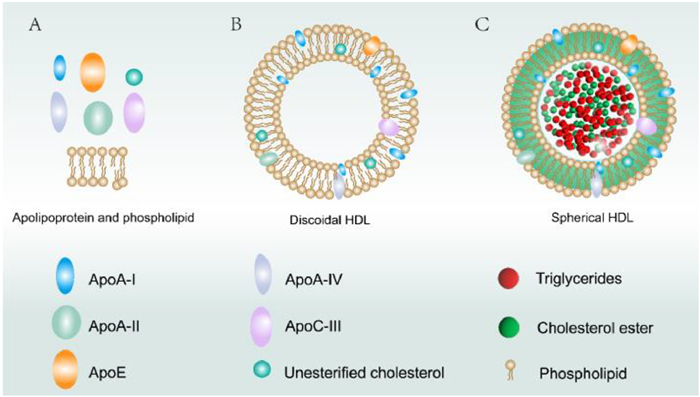
HDLs are complex nanoparticles with specific structures. Understanding their molecular structure and dynamics is crucial to reveal the mechanisms and regulatory principles of their physiological functions. Typically, different HDL nanoparticles exhibit distinct dynamic behaviors based on variations in their compositional components and ratios of each component. The interaction patterns between the different HDL subtypes determine the dynamic characteristics of their constituent parts, including the flexibility of lipids and proteins [18]. HDL consists of two main components (Fig. 1): lipids and proteins [19]. The lipid components include cholesterol, phospholipids, and triglycerides, whereas the protein components include apolipoproteins, lecithin-cholesterol acyltransferase (LCAT), and cholesteryl ester transfer protein (CETP). The first aspect to explore is the distinction between the different compositional components of HDL and the interaction patterns between the different HDL subtypes.
Figure 1
Figure 1.
Schematic representation of the structure of high-density lipoprotein. (A) Some apolipoproteins, including ApoA-I, ApoA-II, ApoA-IV, ApoC-III, ApoE, and unesterified cholesterol, phospholipids. (B) Discoidal high-density lipoprotein. (C) Spherical high-density lipoprotein.
HDLs are rich in lipids, with cholesterol being the main component. Cholesterol present in HDL particles mainly originates from peripheral tissues and cells and is collected from these tissues through the process of reverse cholesterol transport [20]. In addition to cholesterol, HDLs contain phospholipids, of which sphingomyelin (SM) is the major phospholipids. Sphingomyelin and cholesterol levels are closely linked, In the 1980s, researchers discovered that the incorporation of SM into cells reduced low-density lipoprotein degradation. This suggests that after participating in cell membrane synthesis, SM can influence the regulation of cell surface low-density lipoprotein receptors and intracellular cholesterol balance, thereby increasing cholesterol synthesis [21]. It is worth mentioning that recently, Li et al. re-evaluated the interaction between SM and cholesterol in cell membranes and circulation. The ratio of SM to cholesterol directly affects the potential of HDL for cholesterol transport. Additionally, in mice, the overexpression of SM increases atherosclerosis-prone lipoproteins rich in SM and synergistically promotes the development of atherosclerosis along with cholesterol [22]. Apart from cholesterol and SM, HDL also contains over a dozen other lipids, such as phosphatidylcholine and phosphatidylethanolamine [23]. These phospholipids can form lipid complexes with cholesterol and collectively participate in the reverse transport of cholesterol via HDL.
2.2
Protein composition of HDLs
HDL proteins also have crucial structural features. There are various proteins present on HDL particles, the most important being apolipoproteins such as apolipoprotein A-I (ApoA-I). ApoA-I is the most abundant protein in HDL particles, constituting > 70% of its total mass [24]. It plays a significant role in reverse cholesterol transport by binding to free cholesterol and phospholipids, forming HDL-cholesterol complexes, and mediating the efflux of these substances from within cells through transport proteins such as ATP-binding cassette transporter A1 (ABCA1) [20]. ApoA-I also performs other functions in the body, in addition to its role in reverse cholesterol transport. According to the literature, ApoA-I functions as an anti-inflammatory mediator by upregulating membrane-associated protein A1 (annexin A1) in endothelial cells and activating related signaling pathways to prevent endothelial cell inflammation. This enriches our understanding of the anti-inflammatory mechanisms of ApoA-I and further elucidates its protective role in atherosclerosis [25]. Based on the difficulty in extracting ApoA-I from plasma, as well as its susceptibility to oxidation and contamination, a laboratory led by Mendez designed novel ApoA-I mimetic peptides. These amphipathic helical peptides comprise 18 amino acids and can remove cellular cholesterol and phospholipids or interact with binding sites on HDLs. Moreover, the synergistic effect of two or more helical repeat sequences enhances these capabilities [26]. Subsequently, an increasing number of ApoA-I mimetic peptides have been developed for disease treatment. Recently, Delk et al. conducted a comprehensive review on the relationship between apolipoprotein mimetics and tumors [27]. They extensively discussed how mimetic peptides effectively reduced tumor burden and inflammatory lipid signals in animal models of cancer, providing significant insights for further research in this field. In addition, HDL contains several other proteins [23], including apolipoproteins A-II (ApoA-II), A-IV (ApoA-IV), C (ApoC), and D (ApoD). These apolipoproteins function synergistically on HDL nanoparticles and participate in various physiological processes such as cholesterol metabolism, lipid transport, inflammation regulation, and antioxidant activities. Their presence and function are crucial for maintaining an overall balance between lipid metabolism and cardiovascular health. Furthermore, HDL nanoparticles also carry certain enzymes such as CETP and LCAT, which play important roles in HDL metabolism and functionality.
2.3
Subclasses of HDLs
In the circulation, HDLs exist in different subclasses known to have different sizes and compositions. Based on their density and electrophoretic properties, HDL can be classified into several subclasses, such as HDL2, HDL3, which may be further divided into subclasses HDL2a, HDL2b, HDL3a, and HDL3b. HDL2 is a large, less dense, and relatively lipid-rich subclass of HDL, whereas HDL3 is a small, denser, and relatively protein-rich subclass of HDL [28]. These subclasses exhibit differences in structure and function [29], and they play distinct roles in lipid metabolism and cardiovascular health. Li et al. found that in septic patients, the plasma levels of HDL and HDL2b were significantly decreased (P < 0.01), whereas the levels of HDL3 remained normal (P > 0.05). The decrease in plasma HDL2b levels may be linked to worsening of inflammation and adverse prognosis in sepsis, potentially through its impact on the SOCS1 signaling pathway. This is supported by the observation that in mice with downregulated HDL2b, SOCS1 mRNA levels in the liver and spleen were significantly increased [28]. Interestingly, in a study investigating the role of anti-inflammatory HDL subclasses, researchers collected blood samples from 316 participants and used microfluidic chip electrophoresis to detect the HDL subclasses. In multivariate linear regression analysis, it was found that when the HDL2b level increased, the white blood cell (WBC) count, lymphocyte count, and high-sensitivity C-reactive protein (hs-CRP) levels in the blood decreased correspondingly [30]. This suggests an anti-inflammatory effect associated with elevated HDL2b levels. Although different types of HDL particles share a similar general organization and structure, specific regions of the protein scaffold can undergo conformational changes depending on the particle size or composition. These dynamic changes in protein conformation may act as interaction targets for enzymes, such as LCAT and CETP, resulting in different biological behaviors [31]. Furthermore, relevant studies have reported that changes in maternal HDL subclasses can predict the growth and development of newborns [32]. Certain HDL subclasses can also be used to assess the risk of developing colon cancer [33]. Moreover, in certain cases, the lipid content and functionality of the HDL subclasses can better predict cardiovascular risk than HDL as a whole [34]. Nuclear magnetic resonance spectroscopy can be used to study the HDL subclasses and assess their impact on the long-term survival of patients with peripheral artery disease. Bernhard et al. conducted a 10-year follow-up study of 319 stable patients with peripheral artery disease and cardiovascular risk factors. This study demonstrated that nuclear magnetic resonance measurement of HDL particles and HDL subclasses can identify patients with a hidden high risk of peripheral artery disease [35]. In recent years, several research reports on HDL subclasses have emerged and studies are no longer limited to defining the relationship between HDL subclasses and cardiovascular risk. Based on HDL piggybacking on drugs, further use of the biological properties of HDL subclasses to fully utilize the drug-carrying capacity to enable the drugs to achieve a more pronounced clinical effect may become the focus of future research.
2.4
Biosynthesis and metabolism of HDLs
The biosynthesis and metabolism of HDLs are complex processes. Researchers have been enthusiastic about studying the mechanisms of HDL synthesis in the body [36-38], as understanding these mechanisms can provide guidance for the ex vivo synthesis of HDL particles. In our discussion, we have mainly focused on the classic process of HDL synthesis. In 2001, Neufeld's laboratory used the fusion of green fluorescent protein (GFP) with ABCA1 to detect the intracellular localization and transport of ABCA1. They found that ABCA1 plays a role in the efflux of lipids from the cell surface [39]. Later, researchers discovered that HDL particles are thermodynamically stable molecular assemblies consisting of helical apolipoproteins and phospholipids for transporting cholesterol. The primary mechanism of HDL synthesis involves ABCA1, which facilitates the efflux of phospholipids and cholesterol from the intracellular compartment to ApoA-I or other lipid-poor apolipoproteins [38]. HDL synthesis involves the interaction of apolipoproteins with ABCA1, which results in the formation of discoidal lipid-poor nascent HDL. Subsequently, LCAT facilitates the entry of cholesterol into the hydrophobic core of HDL, leading to the transformation of nascent discoidal HDL into spherical mature HDL. In 1975, Schonfeld et al. used lipids and apolipoproteins to reconstitute HDL in vitro using ultracentrifugation. This experiment was mainly conducted to demonstrate that the poor reactivity of ApoA-I in radioimmunoassays was due to its interaction with lipids or other proteins in the circulation [40]. However, the in vitro synthesis of HDL was successful in later studies and facilitated the research on HDL for drug-carrying purposes. As we understand, the ABCA1 protein belongs to the ABC transporter family, and its structure consists of multiple functional domains, including two nucleotide-binding domains (NBD1 and NBD2) and two transmembrane domains (TMD1 and TMD2). The latter are embedded in the cell membrane to form substrate channels. NBD1 and NBD2 comprise a series of conserved amino acid sequences [41,42]. These sequences contain typical features of the ABC transporter protein family and are capable of binding and hydrolyzing the amino acid residues of ATP to provide energy. The TMD1 and TMD2 domains, on the other hand, are composed of a series of transmembrane α-helices and are commonly expressed in certain membrane proteins, G-proteins, and ion channel proteins. These helices form channels in the cell membrane, allowing substrate transport. In the case of ABCA1, the TMD1 and TMD2 domains play crucial roles in the transport of cholesterol and phospholipids [43].
HDL can be synthesized in the organism. It primarily occurs in the liver and intestines, where hepatocytes synthesize and secrete ApoA-I. As mentioned above, ApoA-I binds to lipid components, such as phospholipids and cholesterol, to form primary HDL particles which enter the circulatory system to undergo remodeling and maturation through interactions with other proteins and enzymes. LCAT plays an important role catalyzing the formation of extremely hydrophobic cholesteryl esters from free cholesterol which gradually accumulate in the center of HDL particles [44]. This process not only increases the density of HDL particles but also enhances their cholesterol transport and metabolic functions. The remodeling of discoidal high-density lipoprotein into spherical high-density lipoprotein is primarily due to the accumulation of cholesterol esters in the core of the HDL particles (Fig. 2). In addition to the action of the LCAT enzyme, the remodeling and maturation of HDL also involve lipid modifications and changes in protein composition. This process requires the involvement of various proteins, such as CETP and phospholipid transfer protein (PLTP) [45]. CETP plays a crucial role in transferring cholesterol esters between HDL, LDL, and very low-density lipoprotein (VLDL). It facilitates the transfer of cholesterol esters from HDL to VLDL, and reciprocally transfers triglycerides from VLDL to HDL. As a result, triglycerides accumulate in HDL, whereas cholesterol esters are transported to VLDL. This change allows for greater reverse transport of extrahepatic cholesterol to the liver for metabolism, taking full advantage of LDL and VLDL receptors on the liver surface. For example, in a Mendelian randomization study, Cupido et al. found that the combined inhibition of CETP and PCSK9 increased the risk of cardiovascular disease. A reduction in CETP levels significantly decreases the effective number of HDL particles, thereby compromising the body's ability to handle cholesterol outside the liver [46]. Finally, HDL completes its clearance and metabolism through interactions with receptors and enzymes as well as participation in other metabolic pathways. Specific receptors on HDL particles, such as SR-B1, recognize and bind to HDL, promoting the uptake and transport of cholesterol and other lipids. In the liver and other tissues, cholesterol and lipids undergo further metabolism and transport, contributing to the overall regulation of lipid metabolism.
Figure 2
Figure 2.
Biogenesis of HDL. Lipid-free ApoA-I is synthesised from the liver and secreted into the circulation, where it combines with phospholipids and cholesterol effluxed from cell membranes via ABCA1 to form discoidal HDL. Cholesterol efflux from cell membranes via ABCG1, LCAT converts cholesterol to hydrophobic cholesteryl esters, which enter the core of discoidal HDL. Discoidal HDL is converted to globular HDL.
HDL particles play various roles in the body, including reverse cholesterol transport, anti-inflammatory and antioxidant activities, protective effects in endothelial cells, and anti-apoptotic activity. Additionally, they are involved in the regulation of immune functions [47]. These functions interact and promote each other. HDLs primarily regulate the lipid content in the plasma through reverse cholesterol transport (RCT), transporting cholesterol from peripheral cells (including adipocytes, macrophages, and endothelial cells) back to the liver for metabolism, a process coined as RCT by Glomset [48]. Burke et al. discovered that aspirin can lower the incidence of cardiovascular diseases [49]. However, this effect was not solely due to the anticoagulant properties of aspirin. Further research has revealed that aspirin serves as a substrate for PON 1, an enzyme-like substance that is widely present in HDL particles. HDL can utilize PON 1 to hydrolyze aspirin and release free salicylic acid, a potent antioxidant. Prolonged use of aspirin may slow the progression of atherosclerosis, possibly because of the beneficial effects of free salicylic acid in scavenging free radicals [50]. Subsequent studies have referred to different subclasses and heterogeneity of HDL particles that may play different roles in reverse cholesterol transport [34,35,51,52]. The function of reverse cholesterol transport by HDL in relation to cardiovascular disease was well studied a decade ago, and very good reviews have been reported [53,54]. Currently, research has mainly focused on the transport capabilities of HDLs and their targeted delivery functions. These functionalities, which scientists have only realized in the 21st century, could also be harnessed for their powerful therapeutic effects. Additionally, the anti-inflammatory and antioxidant properties of HDL should not be underestimated because they work in coordination. One of the mechanisms involves HDL and ApoA-I directly binding to and neutralizing lipopolysaccharides or lipoteichoic acid to suppress inflammation [55]. Another mechanism involves the reduction of Toll-like receptor 4 (TLR4) surface expression by inducing cholesterol efflux. This leads to the decreased activation of downstream MyD88 and TRIF signaling pathways, inhibiting nuclear factor kappa B (NF-κB) and type I interferon (IFN) responses [56,57]. These mechanisms work together to endow HDL with anti-inflammatory and antioxidant properties. Furthermore, various components of HDL influence its anti-inflammatory capacity. For example, changes in phospholipid composition can alter the anti-inflammatory activity of HDL. Specifically, an increase in the phosphatidylserine content enhances HDL's anti-inflammatory capability [58]. Moreover, because of the intricate and diverse composition of HDL, its components are constantly subjected to changes influenced by various enzymes and substrates. It remains unclear which specific components of HDL particles, carrying a myriad of proteins and lipids, are responsible for controlling anti-inflammatory activity in endothelial cells. Substantial experimental data are required to establish this conclusively. Currently, the anti-inflammatory capacity of HDL has been shown to potentially provide independent and reliable clinical information for risk assessment in patients with cardiovascular disease (CVD) who may not exhibit initial clinical symptoms. The anti-inflammatory activity of HDL may reflect the underlying pathological and physiological processes associated with atherosclerotic CVD [59]. In summary, the functions of HDL and cardiovascular disease events are closely related. While we study the research progress of HDL drug carrying, we provide a brief description of the long-term research focus of HDL so that researchers can quickly grasp the biological properties of HDL in subsequent studies and continue to explore new research directions for HDL.
4.
Principles and advantages of rHDL as a drug carrier
4.1
Fundamentals and targeting mechanisms of rHDL as a drug delivery system
HDL serves as a lipid carrier in the human body, and esterified cholesterol, owing to its extremely hydrophobic nature, enters the HDL core. Similarly, many chemotherapeutic drugs such as doxorubicin, paclitaxel, docetaxel, lapatinib, cisplatin, and curcumin are also hydrophobic [60]. This results in the inability of the drug to effectively enter tumor tissue and requires either binding to proteins to increase solubility or employing a carrier, such as the natural carrier HDL, to overcome this challenge [61]. Researchers have identified two main mechanisms by which HDLs precisely deliver drugs to specific tissues one of which is the body's own production of HDL in the plasma through interactions with HDL receptors (including ABCA1, ABCG1, and SR-B1), which are often highly expressed in hepatocytes and small intestinal mucosal cells [62] and are mainly involved in cholesterol and phospholipid synthesis, transport, and secretion. In addition, ABCA1 and ABCG1 are highly expressed in brain tissue, adipocytes, and immune cells, where they regulate intracellular lipid transport and maintain cholesterol homeostasis [39,63]. In contrast, SR-B1 is mainly expressed in hepatocytes, which is closely related to the RCT pathway and is a key receptor for the major role of HDL in humans [48,64]. These receptors transport lipids, proteins, and nucleic acids from donor cells to recipient cells, which means that drugs transported by HDL can be precisely transported to target tissues and cells through specific ligand-receptor binding. Based on this targeting mechanism, it has been observed that in certain disease states, damaged tissues may exhibit elevated expression of the SR-B1 receptor. For example, in patients with aldosterone-producing adenoma (APA), the adrenal glands produce excessive aldosterone, the precursor of which is cholesterol. Harashima et al. demonstrated that both SR-B1-mediated selective cholesterol ester uptake and de novo cholesterol synthesis are increased in APA [65]. In cancer, tumor cells exhibit increased active cholesterol metabolism and increased SR-B1 expression. For instance, in patients with adrenocortical carcinoma, the CLA-1 gene plays a role in adrenal steroidogenesis and may act as an HDL receptor, mediating the increased SR-B1 selective uptake of cholesterol in these cells [66]. This conclusion has been validated in other cancers such as breast cancer [67], renal clear cell carcinoma [68], and prostate cancer [69]. Interestingly, tumor tissues exhibit a unique blood vessel structure and abnormal function, which increase the permeability of drugs and facilitate their retention in the tumor tissue. This phenomenon represents a significant difference between tumor tissue and normal tissue, and was first coined as the "enhanced permeability and retention (EPR) effect" of tumors by Professor Maeda in 1986 [70]. This serves as another target mechanism for HDL as a drug delivery system. Notably, after 37 years of research, the existence of this effect has provided a theoretical basis for nanoparticles to serve as drug carriers, as the range of particle diameters allowed to pass by the EPR effect is relatively wide, usually within a few hundred nanometers [71,72]. Nanoparticles in this range can enter tumor tissue through the walls of tumor blood vessels and remain in the tissue for a longer period. Smaller nanoparticles (approximately 10–100 nm) can enter the tumor tissue through gaps in the tumor blood vessels and irregular endothelial cell connections [73]. Owing to the increased permeability of the tumor blood vessel walls, these small nanoparticles can effectively penetrate the tumor tissue. The diameter of the HDL particles is typically between 8 nm and 13 nm, and for reconstituted HDL produced synthetically with different drug loads, the diameter can range from to 9–20 nanometers [74]. Therefore, reconstituted HDL passively targets tumor tissues via the EPR effect. Previous studies have reported that larger nanoparticles (approximately 100–200 nm), such as liposomal nanoparticles, primarily enter the tumor tissue through the lymphatic system. Owing to the abnormal structure and function of the lymphatic system in tumor tissue, these nanoparticles are difficult to effectively clear and tend to remain in the tissue for a longer time, leading to their accumulation in the tumor tissue [75]. It is important to note that particle size is not the only factor that determines its effect through EPR. Other factors such as the surface properties, shape, and surface modifications of the particles, as well as the variable structure of the tumor vasculature, also affect the permeability and residence time of the nanoparticles in the tumor tissue [76,77].
4.2
Advantages and disadvantages of rHDL as a drug delivery system
As many normal organs in the human body, such as the liver, adrenal cortex, testes, and ovaries, require cholesterol for their functions, there is still a demand for cholesterol intake from the plasma, despite the fact that most cholesterol is synthesized within cells [78]. A means of modifying HDL as a drug carrier to achieve precise targeting and accumulation in tumor tissues while preventing HDL from participating in reverse cholesterol transport and accumulating in normal liver tissues, leading to hepatotoxicity is a problem that has frustrated scientists. For this reason, LDL has rarely been used as a drug carrier in recent years. LDL is mainly involved in the transport of cholesterol from the liver to the periphery, and LDL receptors are expressed in many peripheral organs. Although LDL is considered a "bad apolipoprotein" in the cardiovascular system, it is one of the most important sources of cholesterol in other tissues and organs, such as the adrenal cortex, the testes, and the ovaries, to ensure that adequate steroid hormones can be synthesized [79]. For many years, to avoid the hepatotoxicity of HDL-loaded drugs, many basic studies have focused on ways to increase the affinity of mimetic HDL and scavenger receptors that are highly expressed in tumors. For example, in a study using dual-functional biomimetic HDL nanoparticles for apoptosis and antitumor therapy, researchers monitored the DiR fluorescence signal and intensity distribution in nude mice injected with the tested formulations of regular lipid (LNP) loaded with drugs and cell-penetrating peptide (CPP)-anchored recombinant high-density lipoprotein (cp-rHDL) loaded with drugs at 2, 8, and 36 h [80]. They found that the fluorescence signal of LNP/DiR appeared mainly in the liver and not in tumor cells at 2 h, indicating that scavenger receptor-mediated lipid uptake resulted in drug release in the liver, leading to hepatotoxicity. In addition, there was almost no signal indicating rapid lipid clearance after 36 h [80]. In contrast, cp-rHDL/DiR exhibited a significant fluorescence signal at the tumor site 8 h after injection. This biomimetic HDL nanocarrier combined biological stability, biocompatibility, degradability, and receptor affinity with pH-controlled penetration properties, allowing cp-rHDL to achieve optimal cellular uptake and efficient internalization. It effectively delivered the cargo, gambogic acid (GA), to cells without significant cytotoxicity [80]. This finding suggests that HDLs have significant potential for use as drug-carrying systems.
To date, various nanocarrier platforms have been developed for disease treatment [81], including cell membrane-camouflaged drug delivery systems [82], lipids, inorganic materials, polymers [83,84], and extracellular vesicles [85] as biological nanocarriers. A significant difference between the diameters of HDL and liposomes was observed. Liposomes typically have diameters ranging from 50 nm to 200 nm [86], while HDL diameters range from 8 nm to 13 nm. Artificially synthesized HDLs can have diameters of 9–20 nm, depending on drug loading [74]. Studies have shown that in tumor cells, the extracellular matrix has small gaps, with closely packed gaps ranging from 20 nm to 42 nm [87]. Larger nanoparticles may have difficulty penetrating these gaps, but appropriately sized HDL may efficiently penetrate or diffuse into tumor tissues. Moreover, some studies have reported that unmodified liposomes can rapidly bind to plasma proteins or interact with factors such as lysozyme in the blood, leading to their clearance from the body. However, when liposomes are surface-modified with polyethylene glycol (PEG), the circulation time of the drugs can be extended [88]. This problem does not occur in naturally occurring human HDLs.
Inorganic nanocarriers can be further categorized into different types such as metal-based nanocarriers, silica-based nanocarriers, carbon nanotubes, and graphene oxide nanoparticles [89]. Compared to HDL, traditional inorganic nanocarriers lack specific receptor targets. However, they have advantages such as metal-based nanocarriers whose metabolic pathways can be controlled in the body by applying an external magnetic field [90]. Some studies have utilized the biocompatibility of HDL and the "magnetic targeting" of SPIONs to design a hybrid nanocarrier called rHDL-SPION-valrubicin for the treatment of prostate cancer. The results demonstrated that the therapeutic effect of the rHDL-valrubicin-SPIONs was seven times greater than that of free valrubicin alone [13].
Extracellular vesicles, also known as exosomes, are another potential drug carrier. Exosomes are large extracellular vesicles that range from 40 nm to 160 nm in size and are secreted by living cells. They possess high biocompatibility, low toxicity, low immunogenicity, and the ability to cross natural barriers, making them a research hotspot [85]. However, there are many uncertainties regarding the use of exosomes as drug carriers. For example, drug loading may lead to leakage and premature release [91], and upon entering the bloodstream, exosomes are prone to recognition and clearance by the mononuclear phagocyte system (MPS) [92]. Furthermore, the targeting mechanisms of exosomes in the body require further investigation [93]. In summary, compared to other nanomaterials used for drug delivery, recombinant high-density lipoproteins (rHDLs) possess several advantages that make them highly promising for molecular delivery. First, the amphiphilic nature of rHDLs allows them to carry small hydrophobic and hydrophilic molecules, drugs, and proteins. Hydrophobic drugs can enter the core of HDL, whereas hydrophilic drugs can bind to the hydrophilic HDL surface. This characteristic allows HDL to carry multiple drugs, enabling synergistic effects and providing endless possibilities for drug penetration into tumor tissues. Second, high-density lipoproteins have a natural structure that provides biocompatibility with the body. They are less likely to be recognized and cleared by the human reticuloendothelial system (RES), resulting in a longer circulation half-life and inherent targeting properties to different receptor cells. Third, most receptors for HDL expressed by cells in the human body are well known, which enables a better understanding and control of HDL metabolic pathways in vivo, thus providing the potential for targeted drug delivery to cells expressing these receptors.
5.
Preparation of HDL nanoparticles
After years of research, the specific synthesis and metabolism of HDL in the body are well understood, and the preceding sections have described the potential of HDL as a nanocarrier platform. Therefore, it is particularly important to find methods for industrial-scale production of HDLs that mimic the functions of the human body. Some inorganic nanoparticles, such as copper nanoparticles, can now be synthesized using a one-step coordination self-assembly method [94]. Exosome, on the other hand, can be isolated and purified from specific cells using differential centrifugation methods [95]. Through continuous experimentation, several methods for HDL synthesizing HDL have been developed, including isolation from plasma, thin-film hydration method and microfluidic assembly method. Specific steps are shown in Supporting information. Schematic of the synthesis steps are shown in Fig. S1 (Supporting information).
6.
Application of HDL as a drug carrier
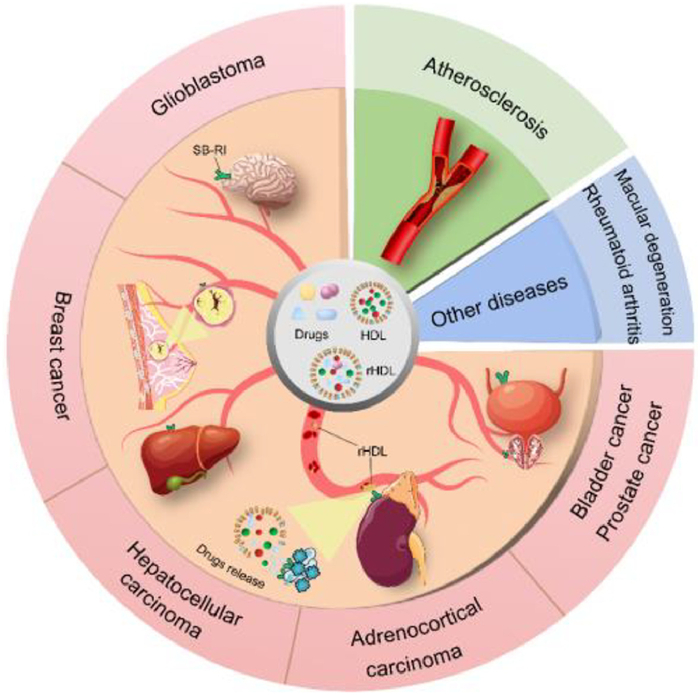
The use of medications for cancer treatment is a popular research topic. The most commonly used chemotherapeutic drugs face several challenges. These include the low water solubility of chemotherapeutic drugs, leading to the need for increased drug dosages in treatment [96]; rapid recognition and clearance of drugs by the mononuclear phagocyte system in the body [97]; insufficient drug accumulation in tumor cells; potential accumulation in other normal tissues causing adverse reactions; and off-target toxicity after repeated drug administration [98]. As previously mentioned, HDL as a drug delivery system possesses targeted effects and is suitable for carrying various hydrophobic and hydrophilic drugs. Current research on drug loading and treatment using HDL has yielded promising results. Additionally, before 2013, several recombinant high-density lipoprotein drugs underwent in clinical trials [99]. Therefore, we gathered research articles published from 2013 to 2023 on the use of recombinant high-density lipoprotein for drug delivery (Fig. 3). Here, we discuss the methodologies and experimental outcomes of these studies and summarize in a table to gain a better understanding of the potential of HDL as a drug carrier for clinical applications (Table S1 in Supporting information).
Figure 3
Figure 3.
The applications of nanomaterial HDL as a drug carrier in cardiovascular diseases, glioblastoma, breast cancer, hepatocellular carcinoma urologic cancers, rheumatoid arthritis and age-related macular degeneration.
The application of HDL in the cardiovascular system was a focus of early HDL research. Studies have shown that synthetic HDL can exert functions such as RCT [100] and the delivery of contrast and therapeutic agents to atherosclerotic plaques [101-103]. In recent years, the trend in using recombinant high-density lipoproteins for anti-atherosclerosis therapy has been moving towards hybridization with polymers, addition of modifications for sustained-release effects, and co-delivery of multiple drugs. For instance, in 2013, a high-density lipoprotein nano-particle was synthesized using an ApoA-I mimetic peptide (residues 221–241) and phospholipids, demonstrating its action in vitro (human and mouse plasma) and in vivo (mice). In this study, the ApoA-I mimetic peptide was altered to 21 amino acids to create a multivalent peptide. This nano-particle exhibited enhanced cholesterol efflux capacity compared to regular recombinant high-density lipoprotein. Upon further observation, it was discovered that the half maximal effective concentration (EC50) value of the modified multimer for cholesterol efflux followed the trend tetramer16 > trimer16 > dimer16, which was proportional to the number of helices in the peptide construct [104]. Convincingly, when the trimeric peptide construct nano-particles were injected into atherosclerotic mouse models during the experimental process, their plasma total cholesterol levels significantly decreased by 50% compared to the control group [104]. In another study, Marrache et al. synthesized TPP-HDL-apoA-I-QD nano-particles that carried diagnostically active quantum dots (QD) and triphenylphosphonium (TPP) cations for imaging and therapeutic purposes against atherosclerosis. The experimental results demonstrated that the stability, immunogenicity, and in vivo retention time of the recombinant high-density lipoprotein nano-particles were satisfactory [102]. The TPP cations in the nano-particles could target the mitochondrial membrane potential (Δψm) of smooth muscle cells and macrophages, enabling the monitoring of apoptosis in these cells. The experiments revealed that, compared to healthy cells, apoptotic macrophages treated with these nano-particles showed a significant reduction in TPP cation accumulation during the early induction of cell apoptosis, as the mitochondrial gradient was lost. Moreover, the injection of TPP-HDL-apoA-I-QD nano-particles in mice resulted in a remarkable decrease in serum total cholesterol and triglyceride levels compared with those in the saline control [102]. rHDL plays multiple roles in atherosclerotic plaque imaging and treatment. Observing the apoptotic state of cells more intuitively and clearly during the treatment process, thus creating a good feedback process, is a promising research strategy.
6.1.2
rHDL loaded with anti-inflammatory drugs
In addition to its inherent role in reverse cholesterol transport and anti-inflammatory effects against atherosclerosis, HDL can also act as a carrier for other anti-inflammatory drugs, leading to synergistic effects. This may produce better results than using nanomaterials alone to piggyback drugs to treat atherosclerosis [15,105]. Statins exhibit significant anti-inflammatory and LDL-antagonistic properties. However, their low bioavailability and potential adverse effects on muscles and liver at high doses are concerning. Nanotechnology has emerged as a solution to improve the bioavailability of statin drugs [106]. For instance, [S]-rHDL nanoparticle carrying simvastatin and amphiphilic magnetic resonance imaging (MRI) contrast agents (gadolinium–diethylene triamine pentaacetic acid–distearyl amide and fluorescent dyes) have been synthesized using a thin film hydration method [103]. In vitro experiments were conducted to assess the effects of [S]-rHDL on hepatocytes, endothelial cells, and smooth muscle cells. Compared with the control group treated with rHDL alone, the viability of various cells was almost completely compromised after 48 h of exposure to [S]-rHDL [103]. Subsequent in vivo studies in mice involved long-term low- and high-dose [S]-rHDL infusions. These results indicated that [S]-rHDL exhibited effective local anti-inflammatory effects in atherosclerotic plaques. This effect may be attributed to a reduction in inflammatory protease activity and inhibition of the mevalonate pathway in mice [103]. It is worth noting that the side effects associated with high-dose simvastatin are not acceptable for human use. Therefore, the long-term efficacy of low-dose [S]-rHDL in mice is crucial. In subsequent studies, poly(lactic-co-glycolic acid) (PLGA) was incorporated to improve the in vivo pharmacokinetics of statins. PLGA-HDL nanoparticles can facilitate controlled drug release and exhibit significant accumulation in atherosclerotic plaques [107]. Furthermore, studies have indicated that the cell adhesion molecule, CD44, is upregulated in atherosclerotic plaques and damaged endothelial cells in certain tumors. Hyaluronic acid (HA) serves as a major endogenous ligand of CD44 [108]. Therefore, HA-modified rHDL accumulate more accurately in atherosclerotic plaques than in the liver. Liu et al. utilized hyaluronic acid (HA) to modify rHDL loaded with lovastatin (LT), resulting in HA-LT-rHDL [101]. The hepatic uptake of HA-LT-rHDL was significantly reduced to approximately 1.61 times lower compared to that of LT-rHDL (Fig. S2 in Supporting information). Owing to the targeted binding of HA to CD44, the accumulation of HA-LT-rHDL within atherosclerotic plaques was more than six times that of LT-rHDL [101]. Three years later, the research team refined the application of rHDL for anti-atherosclerotic therapy for drug delivery in atherosclerosis using a combination of PLGA and HA to modify rHDL and loaded it with simvastatin to synthesize HA-(C)-PLGA-rHDL [109]. The experimental results demonstrated its superior efficacy in animal models compared to other formulations for combating atherosclerosis. In addition to the statins commonly used to treat atherosclerosis, certain polymers targeting microRNA-155 (miR155) have been designed to specifically bind to miR155 expressed in smooth muscle cells within atherosclerotic plaques. This inhibition effectively blocks the transfer of exosome-mediated miR155 between smooth muscle and endothelial cells, reducing detrimental lipid metabolic events in the cardiovascular system [110]. Lu et al. reported a study using recombinant high-density lipoprotein nanoparticles loaded with anti-miR155 for combined anti-atherosclerosis therapy at the cellular level. The experimental results showed that both the anti-miR155 and recombinant high-density lipoprotein inhibited the cellular uptake of oxidized LDL (oxLDL) and reduced the generation of reactive oxygen species, thereby mitigating foam cell formation [111]. Unfortunately, this nanoparticle, equipped with anti-miR155, has not been studied in animal models of atherosclerosis, and more in-depth experiments are needed for validation.
Despite the satisfactory results obtained from previous studies using HA to enhance the targeting ability and PLGA to control drug release, repeated administration still induces accelerated drug clearance in vivo. Researchers have discovered that ganglioside GM1 modification can increase the stability of liposomes, prolong circulation time, and reduce clearance by the reticuloendothelial system [112]. Consequently, GM1-modified rHDL loaded with lovastatin (LT-GM1rHDL) was designed using the thin-film hydration method described by Wang et al. [113]. After administering LT-GM1rHDL to animal models, the drug concentration remained at very low levels even after 36 h, in stark contrast to the LT solution at 12 h, and LT-loaded nanostructured lipid carriers and LT-rHDL (used for comparison) at 24 h in various control groups [113]. This may be because the specific recognition of GM1 oligosaccharides by the reticuloendothelial system slows down during the metabolic process, resulting in a prolonged circulation time of GM1-modified rHDL [113]. Moreover, a recent study using rHDL loaded with GM3 for the treatment of atherosclerosis found that exogenous GM3 molecules, through pH-responsive release, induced the removal of other major lipids (such as cholesterol and phospholipids) after entering atherosclerotic plaques [114]. This pH-responsive release effect may potentially reduce the dosage of exogenous drugs, thus minimizing the side effects. For example, pH-sensitive nanomaterials can improve the delivery efficiency of anti-tumor drugs [115]. In a recent study, Li et al. have designed a hydrogel that achieves sustained drug release at pH 5.8 to improve the anti-tumor efficacy in vivo [116]. However, this promising speculation that requires extensive experimental evidence to confirm its accuracy.
6.1.3
rHDL loaded with other drugs
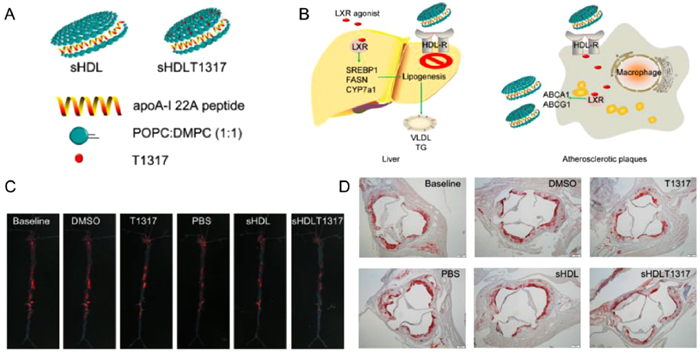
In addition, rHDLs have been loaded with other formulations for the treatment of atherosclerosis. Guo et al. designed rHDL-carrying liver X receptor (LXRs) agonists to promote cholesterol efflux from macrophages for the treatment of atherosclerosis [117]. When LXR-rHDL was injected into ApoE-deficient mice fed a high-fat and high-cholesterol diet, LXRs were targeted atherosclerotic plaques and activated LXR target gene expression (Fig. 4). This led to cholesterol efflux from plaques and facilitated the RCT pathway, thus exerting therapeutic effects [117]. In another study, Rink et al. synthesized a vascular disease-targeting biomimetic nanotherapeutic that combined the functions of nitric oxide (NO) and HDL, called S-nitrosylated (SNO) phospholipid (DPPNOTE), taking advantage of the ability of NO to dilate blood vessels and act as a cellular second messenger to attenuate endothelial damage [118]. It has also been demonstrated that SNO HDL NPs inhibit nuclear factor-κB (NF-κB) activation through the sustained release of NO, thereby resisting inflammation, promoting cholesterol efflux, inhibiting the migration of aortic smooth muscle cells, decreasing ischemia/reperfusion injury, and decreasing atherosclerosis, among other beneficial effects on vascular diseases [118].
Figure 4
Figure 4.
Promoting atherosclerotic regression through high-density lipoprotein-targeted delivery of hepatic X-receptor agonists. (A) Schematic structure of T1317-encapsulated sHDL NPs. (B) Schematic diagram of the principle mechanism of drug delivery targeting by sHDL NPs. (C) whole aortas excised for plaque area analysis by oil red O staining. sHDLT1317-treated mice exhibited decreased oil red O-stained lesions in the whole aorta. (D) The reduction in aortic root plaque area was most pronounced in sHDLT1317-treated mice. Reproduced with permission [117]. Copyright 2018, Elsevier.
In summary, research into rHDL-based drug delivery for the treatment of atherosclerosis is ongoing. Currently, an increasing number of anti-atherosclerotic drugs are being combined with HDL to maximize their therapeutic effects. rHDLs hold significant research prospects for clinical application in conditions affecting the cardiovascular system.
6.2
Cancer
6.2.1
Glioblastoma
The nervous system is often not easily accessible to drugs used to treat brain disorders because of the presence of the blood-brain barrier (BBB), which prevents drugs entering brain tissue. Therefore, researchers are in dire need of developing methods to overcome the limitations of the BBB, so that drugs can better treat certain neurological disorders [119]. SR-BI is expressed on the endothelial cells of brain capillaries, and HDLs recognize SR-BI receptors and freely cross the blood-brain barrier [120]. Therefore, it becomes possible for drugs to piggyback on rHDLs to treat neurological disorders. As early as 2013, researchers found that HDLs exhibited varying affinities for brain SR-BI receptors under different conditions. Huang et al. constructed discoidal (d-rHDL) and spherical (s-rHDL) rHDLs using apolipoprotein E and compared their efficiency in targeted delivery to glioblastoma (GBM) [121]. They observed that after injecting both types of rHDLs, s-rHDLs consistently displayed higher fluorescence intensity at various time points at the tumor site. Compared to d-rHDL, s-rHDL showed a 73% increase in fluorescence intensity, indicating that s-rHDL penetrated the tumor microenvironment of GBM [121]. Based on the fact that glioblastoma is a typical Ras-activated cancer and the macroautophagy pathway, which is almost absent in normal cells, is highly activated in Ras-activated cancer cells [122], targeting the macroautophagy pathway offers a new therapeutic approach for treating refractory glioblastoma. In light of this, Huang et al. designed rHDL loaded with calcium phosphate (CaP)-encapsulated siRNA that could be precisely delivered to Ras-activated glioblastoma cells through the process of macroautophagy [123]. This targeting mechanism differs from receptor recognition-based targeting and can circumvent the side effects resulting from the recognition of loaded drug-rHDL by receptors in peripheral normal tissues (Fig. S3 in Supporting information). However, drug delivery for treating glioblastoma using rHDLs is not only influenced by the BBB but also by the blood-brain tumor barrier (BBTB). Enhancing targeted drug delivery can reduce drug dosage and minimize the occurrence of side effects. Research has shown that the T7 peptide has a high affinity for transferrin receptors (TfRs) overexpressed in glioblastoma, whereas the dA7R peptide has a high affinity for vascular endothelial growth factor receptor 2 (VEGFR2), which is highly expressed in brain glioblastoma cells [124]. In order to improve the efficiency of drug delivery to glioblastomas, Cui et al. used T7 and dA7R peptides to enhance the targeting of HDL, actively targeting glioblastoma cells by interacting with TfR and VEGFR2, thereby overcoming both the BBB and BBTB [124]. In the experiments, HDL modified with T7 and dA7R peptides loaded with the hydrophobic anticancer drug 10-hydroxycamptothecin demonstrated excellent targeting specificity. In a mouse model, more Cy5.5-labeled T7/dA7R-HDL accumulated in glioblastomas, and its therapeutic efficacy in mice was not inferior to the control group [124]. In another study, researchers developed rHDL-based immunotherapy to induce glioblastoma regression. They loaded rHDL nanodiscs with a Toll-like receptor 9 agonist (CpG) and tumor-specific neoantigens, which could induce more effective cytotoxic CD8a+ T lymphocytes in the body, enhancing the body's antitumor response. Experimental results have shown that tumor-specific neoantigens can trigger effective antitumor immunity in a mouse glioblastoma model and promote long-lasting immune memory to prevent tumor recurrence [125].
The application of rHDLs in the nervous system is not limited to tumors; studies have shown that the preparation of relevant nanoparticles can also treat Alzheimer's disease [126,127], multiple sclerosis [128], and C-type Niemann-Pick's disease [129]. In conclusion, the use of rHDLs to enhance BBB penetration is a promising approach for the treatment of nervous system diseases. In addition to gliomas, other Ras-activated tumors may also be suitable for the effective targeted delivery of therapeutic agents to tumor tissues through macropinocytosis, but this needs to be verified by further experiments.
6.2.2
Breast cancer
Cholesterol metabolism plays a crucial role in breast cancer cell growth and migration. Studies have shown that HDL cholesterol and SR-BI are involved in regulating cholesterol homeostasis in breast cancer cells [130]. Therefore, utilizing the SR-BI-mediated cellular uptake mechanism allows the direct delivery of therapeutic drugs to the cytoplasm of tumor cells, achieving drug accumulation [131]. For instance, small interfering RNA (siRNA) is a promising gene therapy that targets genes that promote tumor growth, catalyzes the degradation of already transcribed mRNA, and achieves tumor regression. However, siRNAs are susceptible to degradation by nucleases in the bloodstream and are rapidly cleared by lysosomes even if they enter the cytoplasm of tumor cells [132]. To address this, siRNA delivery systems mainly employ nanocarriers, such as liposomes [133,134], exosomes [135], iron oxyhydroxide (FeOOH) nanoparticle (NP) [136], and HDL nanoparticles [131]. Research has demonstrated the enhanced delivery capabilities of HDL, for instance, rHDLs can carry an siRNA targeting the vascular endothelial growth factor (VEGF) gene, thereby inhibiting tumor angiogenesis [131]. In a nude mouse model, intravenous injection of 25 mg of rHDL/Chol-siRNA-VEGF decreased tumor volume to 54.5% of its original volume after 24 days. At a higher dose (50 mg), the tumor volume decreased to 30.7%. However, intravenous injection of 50 mg of Chol-siRNA-VEGF alone hardly suppressed tumor growth [131]. Subsequently, VEGF expression levels were assessed using Western blot and enzyme-linked immunosorbent assay (ELISA). The results indicated that the mechanism underlying the reduction in tumor volume was a significant decrease in relative VEGF protein expression levels [131]. The utilization of rHDL maximizes the effect of siRNA at the genetic level for the treatment of breast cancer. In addition to delivering siRNA for breast cancer treatment, traditional anticancer drugs such as paclitaxel (PTX) can be combined with rHDL and other novel agents to treat PTX-resistant breast cancer. Zhang et al. utilized rHDL to load two hydrophobic drugs: the PTX and a cancer multidrug resistance reversal agent (HZ08). By monitoring the changes in the concentrations of PTX and HZ08 in the breast cancer cell culture medium, which reflects cellular drug uptake, they found that the PTX-HZ08-rHDL nanoparticles exhibited the highest intracellular accumulation of PTX. This suggests that PTX-HZ08-rHDL nanoparticles may enhance anticancer efficacy delivering PTX to drug-resistant tumor cells through an rHDL-targeting effect and by increasing PTX accumulation with HZ08 [137].
For the treatment of triple-negative breast cancer (TNBC), conventional chemotherapy regimens have shown limited efficacy owing to the lack of well-defined tumor targets, and patients with TNBC typically have the poorest prognosis compared to those with other cancer subtypes. Fortunately, it has been discovered that the sonic hedgehog signaling pathway is highly active in TNBC. The inhibition of this pathway by GANT61 may slow the growth and metastasis of TNBC [138]. Additionally, research has shown that the tLyP-1 peptide is a ligand for the neuropilin-1 (NRP-1) receptor, which is highly expressed on breast cancer cells. Through the specific binding between tLyP-1 and NRP-1, drugs can be delivered to TNBC [139]. In a study aimed at treating TNBC, Jiang et al. used tLyP-1 peptide-modified rHDL to deliver PTX and GANT61, forming tLyP-1-rHDL-PTX/GANT61 nanoparticles. With the dual targeting effect of tLyP-1 and rHDL, these nanoparticles effectively targeted both the NRP-1 and SR-BI receptors on breast cancer cells, facilitating the precise delivery of PTX and GANT61 [140]. In vitro cytotoxicity assays demonstrated that tLyP-1-rHDL-PTX/GANT61 nanoparticles induced higher levels of cell toxicity and death in TNBC cells than free PTX and GANT61 solutions, likely because of the increased accumulation of PTX and GANT61 inside tumor cells [140]. In mouse animal models injected with tLyP-1-rHDL-PTX/GANT61 nanoparticles, enhanced antitumor growth capabilities were observed compared to the control group, and hardly any fluorescent signals indicating metastasis were detected in the mouse lungs (Fig. S4 in Supporting information) [140]. Additionally, rHDL can carry other hydrophobic drugs such as WGA-TA [141], a heat shock protein 90 inhibitor known to induce cancer cell death and possesses potent anticancer activity, to treat TNBC [142].
6.2.3
Hepatocellular carcinoma
In liver cancer cells, the selective uptake of cholesterol mediated by SR-BI is significantly increased, and the RCT pathway shows enhanced activity. Therefore, the use of rHDL to deliver drugs for the treatment of liver cancer has natural advantages. For instance, utilizing rHDL to deliver siRNA into liver cancer cells can knockdown genes that promote cancer cell growth and development, thereby exerting therapeutic effects [143]. In addition to siRNA delivery, hydrophobic drugs such as sorafenib (So) require encapsulation in nanoparticles to achieve better therapeutic efficacy [144]. Sorafenib is an orally administered kinase inhibitor that is widely used in the treatment of liver cancer. However, it has limitations, such as poor solubility, low bioavailability, and susceptibility to elimination, which greatly restrict its clinical application [145]. Furthermore, antimiRNA21, which silences miRNA21, can increase tumor sensitivity to chemotherapy [146]. A combination of So and antimiRNA21-loaded rHDL was designed by Li et al., and it was found that the tumor accumulation of RGD-rHDL/So/antimiRNA21 was 4.2 times higher than that of Lipos/So/antimiRNA21, demonstrating precise targeted delivery of the drugs to the tumor cells [147]. In a mouse model of liver cancer, combination treatment with So and antimiRNA21 using RGD-rHDL/So/antimiRNA21 achieved the highest tumor inhibition rate, reaching 82.13% compared to the control group [147]. This indicates a synergistic effect of the combination therapy, which cannot be achieved using So or antimiRNA21 alone (Fig. S5 in Supporting information). It is worth mentioning that researchers have discovered that liver cancer cell have nuclear localization sequences (NLS), which can be precisely targeted with carbon dots [148]. Could they further enhance the targeting ability of drug carriers when combined with rHDL? This needs to be verified by further experiments.
6.2.4
Urologic cancers
Genitourinary tumors are common malignancies, including adrenal cortical carcinoma, renal clear cell carcinoma, urothelial carcinoma, bladder cancer, prostate cancer, and urethral carcinoma. Currently, the main treatment modalities for these tumors are surgery, radiotherapy, and chemotherapy [149,150]. Over the past decade, research shows that rHDL-based drug delivery has mainly been explored for the treatment of genitourinary tumors, particularly adrenal cortical carcinoma, bladder cancer, and prostate cancer. Therefore, rHDLs have significant potential in the research and treatment of genitourinary tumors.
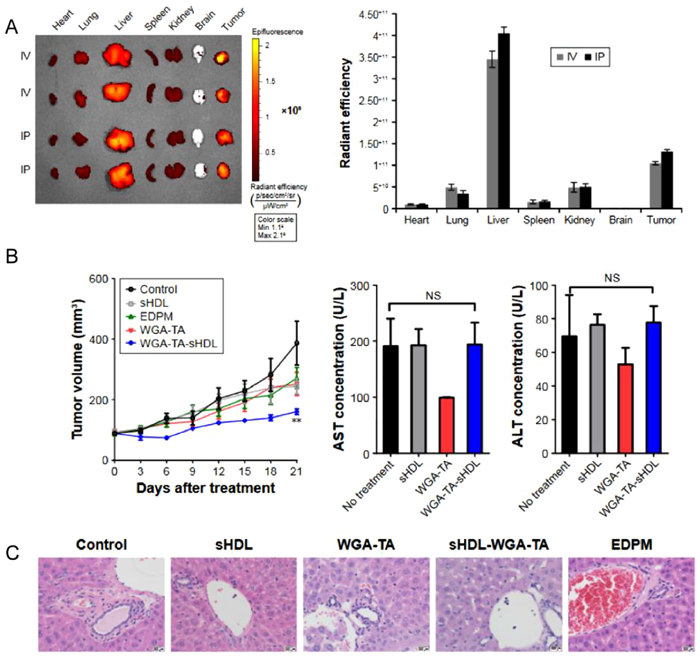
Adrenocortical carcinoma (ACC) is a rare and aggressive malignant tumor of the endocrine system. Clinically, mitotane is commonly used in combination with etoposide, doxorubicin, and cisplatin as first-line treatment for metastatic ACC [151]. However, this combination therapy often result in significant side effects [152]. Subramanian et al. discovered that cholesterol-free sHDL could synergistically enhance the effects of these drugs and reduce the drug concentrations required to achieve the same therapeutic outcome, thereby reducing the incidence of side effects [153]. Compared to a control group receiving monotherapy with chemotherapeutic drugs, the combination of sHDL and chemotherapeutic drugs increased apoptosis by 30%–50% (P < 0.03) and reduced cortisol production by 70%–90% (P < 0.01). This effect may be attributed to the ability of sHDLs to deplete cholesterol in ACC cells through the RCT pathway, thereby preventing the synthesis of steroids necessary for cancer cell growth and development, and leading to the inhibition of cancer cell proliferation [153]. ACC cells overexpress SR-BI to meet the high demand for cholesterol [66]. In another study, s-HDL nanodiscs were loaded with an HSP90 inhibitor (WGA-TA) for targeted delivery to ACC cells [154]. After incubating WGA-TA-sHDL with ACC cell lines expressing high (H295R) and low (SW13) levels of SR-BI for 2 h, over 80% of H295R cells exhibited fluorescent signals, whereas only 10% of SW13 cells showed fluorescent signals. This targeting mechanism, based on SR-BI recognition, facilitates drug accumulation in ACC cells [154]. Although this nanodrug showed fluorescence signals that accumulated in tumors in animal models, it also produced fluorescence signals in the liver, which is the main organ for eliminating nanoparticles (Fig. 5) [154]. Another possibility is that WGA-TA-sHDL is taken up by the liver earlyon, leading to hepatotoxicity. Further experiments are required to verify this hypothesis and improve the targeting capability of sHDL for ACC treatment.
Figure 5
Figure 5.
Delivery of WGA-TA via high-density lipoprotein nanodisks for the treatment of adrenocortical carcinoma. (A) Biodistribution of Dir-shDl in H295 tumor-bearing mice. H295 tumor-bearing mice were injected intravenous or intraperitoneal with DiR-sHDL, and major organs were harvested and imaged 24 h after injection. (B) In vivo therapeutic effect of Wga-Ta-shDl. Mean tumor growth curves and serum levels of AST and ALT in mice using different preparations. (C) After 21 days of treatment, livers treated with different preparations were stained for H & E. Reproduced with permission [154]. Copyright 2017, Informa UK Limited.
Studies have revealed a close association between lipid metabolism and urothelial bladder cancer (UBC). It has been found that ox-LDL increases the number of UBC tumor stem cells through the CD36/JAK2/STAT3 axis, and HDL may counteract these effects and potentially reverse UBC progression [155]. Additionally, abnormal cholesterol metabolism in UBC cells can affect the efficiency of lipid membrane synthesis, stability of lipid energy release, lipid signaling, and activation of inflammation-related pathways [156]. This evidence suggests the relevance of lipid metabolism in UBC, indicating the potential of rHDL for targeted drug delivery to UBC cells. In a study using an rHDL delivery system for treating bladder cancer, the AT1R blocker candesartan (CD) and the p53 gene were delivered to UBC cells using rHDL [12]. Candesartan and p53 exerted synergistic effects by inhibiting tumor angiogenesis. Comparative analysis showed that treatment with rHDL-CD-PEI-p53 complexes in the animal model resulted in significantly smaller tumor volumes (385.07 ± 86.19 mm3) compared to those using CD or p53 alone [12].
Furthermore, extensive research has demonstrated a close association between lipid metabolism and prostate cancer development [157-159]. Elevated transcriptional expression of SR-B1, which is essential for the uptake of cholesterol by cancer cells and the synthesis of steroids and nonsteroidal substances, has been observed in primary prostate cancer. These findings emphasize the potential of targeting SR-B1 through rHDL for drug delivery as a novel approach for the treatment of prostate cancer [160]. For example, siRNA therapy for prostate cancer can be administered using rHDL as a carrier. In an animal model, 13 intravenous injections of rHDL-siRNA resulted in a significant reduction in tumor volume in treated mice [161]. To achieve more precise targeting of prostate cancer cells, Sabnis et al. used magnetic targeting to enhance the drug delivery capacity of lipoproteins. They encapsulated the anticancer drug cabazitaxel and SPIONs into rHDL nanoparticles. Through the combined action of magnetic guidance and rHDL recognition by SR-B1, they maximized cabazitaxel accumulation in cancer cells. The experimental results demonstrated that the rHDL-cabazitaxel-SPIONs exhibited a therapeutic effect seven times greater than that of free cabazitaxel alone (Fig. S6 in Supporting information) [13]. Overall, both the therapeutic and delivery capabilities of rHDL have shown outstanding performance in the treatment of urological tumors.
6.3
Other diseases
In addition to its remarkable drug delivery capabilities in cardiovascular and nervous systems and various cancers, rHDL has shown excellent performance in the treatment of other conditions. In recent studies, rHDL was utilized to deliver xylopic acid [162] and HIF-1α siRNA [163] for the treatment of rheumatoid arthritis (RA), demonstrating precise targeting ability and enhancing the drug's ability to suppress multiple inflammatory signals involved in RA pathogenesis. rHDL have been used to package rapamycin (Rap) to treat age-related macular degeneration (AMD). The incorporation of Rap into s-HDL nanoparticles resulted in a 125-fold increase in its water solubility [164]. In vivo studies have shown that intravitreal administration of s-HDL-Rap in rats accurately reached the retinal pigment epithelium (RPE) layer, significantly reducing lipofuscin accumulation and enhancing the body's anti-inflammatory capacity compared with free drugs or standalone s-HDL treatment [164].
7.
Clinical research
Currently, most research on drug delivery using rHDL nanoparticles is in the preclinical stage, with only a few HDL mimetics for cardiovascular diseases entering clinical trials (Table S2 in Supporting information). We have discussed CER-001 and CSL-112, two rHDL nanoparticles that entered clinical trials. Specific information is shown in Supporting information.
8.
Conclusions and challenges
Over the years, the clinical translational potential of rHDL has been promising. rHDL shares similar biological characteristics with HDL, not only participating in cholesterol metabolism and transport, but also possessing various functions such as antioxidant, anti-inflammatory, and antiplatelet activities. As a drug delivery carrier, rHDL exhibits amphiphilic properties and excellent biocompatibility, along with precise intrinsic targeting, making it advantageous for the delivery of various drugs and molecules. Currently, several methods have been employed to encapsulate hydrophobic substances such as siRNA, drugs, and imaging agents into an rHDL nanoparticle core composed of phospholipids, cholesterol, and proteins, which offer high controllability and stability. These nanoparticles can effectively protect loaded drugs from enzymatic degradation or immune responses in the bloodstream, allowing rHDL to serve as a carrier. Moreover, researchers are continuously refining the characterization and stability assessment methods for rHDL nanoparticles, seeking improved drug loading and release techniques, and exploring the effectiveness and safety of rHDL nanoparticle drug delivery. These studies have yielded promising results in animal models of various diseases. However, despite demonstrating promising drug-loading capabilities in preclinical research, no clinical trials involving rHDL-carrying drugs have been found in the U.S. Clinical Trial Database (www.clinicaltrials.gov). Only a few HDL analogs have been tested in clinical trials. Disappointingly, the clinical trial results of CER-001 were not as ideal as expected and were likely influenced by various factors such as different clinical design conditions, patient quality, and the source of CER-001. Nonetheless, the clinical trial performance of CSL-112 has met expectations, and if it passes phase III clinical trials, it will undoubtedly enhance researchers' confidence in the clinical translational research of rHDL.
However, the challenges faced by rHDLs during drug loading cannot be overlooked. The production process of rHDL is relatively complex and its high cost may limit its large-scale application. Currently, the primary method used in laboratories for rHDL synthesis is thin-film hydration, which cannot be used for large-scale production. Although microfluidic technology can synthesize rHDL in batches, the equipment is expensive and complex, requiring specialized knowledge for implementation which may result in higher production costs. Therefore, developing a standardized method for the large-scale production of rHDL is of milestone significance for rHDL drug loading efforts. Furthermore, we must continue to enhance the targeting effects of rHDL and minimize the side effects of drug delivery. In recent years, the trend in rHDL research has shifted towards hybridization with polymers, adding modifications to achieve sustained release and targeting effects, and loading multiple drugs for synergistic effects. These efforts have enhanced the targeting capabilities of rHDL. Finally, there are differences in the metabolic pathways of humans and model animals. Although rHDL drug loading has demonstrated good biocompatibility in animal studies, considering the potential adverse reactions during long-term and high-dose applications in humans, more clinical trials are required for safety evaluation. Therefore, making rHDL a viable platform for clinical applications is challenging.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Nos. 52273128, 82003450), the Natural Science Foundation of Guangdong Province (No. 2022A1515220048), the Natural Science Foundation of Guizhou Province (Nos. Guizhou Province-ZK [2022] General 300, Guizhou Province-ZK [2023] General 244), the Xiangshan Talented Scientific Research Foundation of Zhuhai People's Hospital (No. 2023XSYC-01) and Guangdong Provincial Key Laboratory of Tumor Interventional Diagnosis and Treatment (No. 2021B1212040004).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2024.109584.
Figure 2
Biogenesis of HDL. Lipid-free ApoA-I is synthesised from the liver and secreted into the circulation, where it combines with phospholipids and cholesterol effluxed from cell membranes via ABCA1 to form discoidal HDL. Cholesterol efflux from cell membranes via ABCG1, LCAT converts cholesterol to hydrophobic cholesteryl esters, which enter the core of discoidal HDL. Discoidal HDL is converted to globular HDL.
Figure 3
The applications of nanomaterial HDL as a drug carrier in cardiovascular diseases, glioblastoma, breast cancer, hepatocellular carcinoma urologic cancers, rheumatoid arthritis and age-related macular degeneration.
Figure 4
Promoting atherosclerotic regression through high-density lipoprotein-targeted delivery of hepatic X-receptor agonists. (A) Schematic structure of T1317-encapsulated sHDL NPs. (B) Schematic diagram of the principle mechanism of drug delivery targeting by sHDL NPs. (C) whole aortas excised for plaque area analysis by oil red O staining. sHDLT1317-treated mice exhibited decreased oil red O-stained lesions in the whole aorta. (D) The reduction in aortic root plaque area was most pronounced in sHDLT1317-treated mice. Reproduced with permission [117]. Copyright 2018, Elsevier.
Figure 5
Delivery of WGA-TA via high-density lipoprotein nanodisks for the treatment of adrenocortical carcinoma. (A) Biodistribution of Dir-shDl in H295 tumor-bearing mice. H295 tumor-bearing mice were injected intravenous or intraperitoneal with DiR-sHDL, and major organs were harvested and imaged 24 h after injection. (B) In vivo therapeutic effect of Wga-Ta-shDl. Mean tumor growth curves and serum levels of AST and ALT in mice using different preparations. (C) After 21 days of treatment, livers treated with different preparations were stained for H & E. Reproduced with permission [154]. Copyright 2017, Informa UK Limited.

 DownLoad:
DownLoad:


 下载:
下载:






 下载:
下载:
