
引用本文:
寻看雨, 孙悦, 张悦, 邱丽萍. 基于功能核酸的细胞荧光成像[J]. 化学通报,
2021, 84(2): 98-107.

Citation: Kanyu Xun, Yue Sun, Yue Zhang, Liping Qiu. Functional Nucleic Acid-Based Fluorescence Cell Imaging[J]. Chemistry, 2021, 84(2): 98-107.
Citation: Kanyu Xun, Yue Sun, Yue Zhang, Liping Qiu. Functional Nucleic Acid-Based Fluorescence Cell Imaging[J]. Chemistry, 2021, 84(2): 98-107.
基于功能核酸的细胞荧光成像
摘要:
细胞是生物体基本的结构和功能单元,对活细胞中特定生物组分进行动态分析,将为相关生命活动过程的研究提供重要信息。荧光成像为细胞分析提供了一种操作简单、灵敏度高、可实时监测细胞微观动态分子过程的光学生物成像技术。发展高性能的荧光探针用于活细胞成像已成为研究热点。功能核酸是一类具有特殊化学和生物学功能的寡核苷酸分子,除了天然存在的核酶(Ribozyme)和核糖开关(Riboswitch)之外,还包括通过指数富集的配体系统进化技术(SELEX)筛选获得的核酸适体和脱氧核酶(DNAzyme)。功能核酸由于具有合成简单、免疫原性低、相对分子质量小、化学稳定性高、易于修饰等优点,在生物成像领域受到广泛关注。本文主要综述了基于功能核酸的荧光探针在细胞成像领域中的应用研究,总结了该领域面临的挑战,并对其未来发展方向进行了展望。
English
Functional Nucleic Acid-Based Fluorescence Cell Imaging
Abstract:
Cell is the structural and functional unit of organisms. Studying the spatiotemporal distribution and activity of specific biomolecules in living cells would provide valuable information in understanding various biological processes. Fluorescence imaging technology has showed great potential for live-cell imaging. On the other hand, development of high-performance fluorescent probes has become a major bottleneck in this field. Functional nucleic acids (FNAs) are a class of oligonucleotides with specific chemical and/or biological functions, including natural ribozymes and riboswitches, as well as aptamers and DNAzymes screened through the SELEX (systematic evolution of ligands by exponential enrichment) technique. Taking advantages of simple synthesis, low immunogenicity, small size, high chemical stability, easy modification, FNAs have received extensive attention in the fields of bio-analysis and bio-imaging. This review summarizes the recent applications of FNA in fluorescence cell imaging, and points out the challenges and future prospects in this field.
-
Key words:
- Functional nucleic acids
- / Fluorescence probe
- / Cell imaging
- / Bioanalysis
-
细胞是生命活动的基本结构和功能单位。生物体的生长、发育、代谢、应激和病变等重大生命现象的研究都以细胞为基础。在活细胞中对重要生物组分如金属离子[1]、小分子[2]、蛋白[3]、核酸[4]等的时空分布及功能活性进行精准测量与调控,将对生命运行规律的探索、疾病发生发展机理的阐明、疾病的预防与临床诊治等具有重要意义。然而,活细胞中生物组分具有种类多、丰度低、分布复杂、动态多变等特点,给活细胞的研究带来了巨大挑战。传统的细胞分析技术(例如western blot[5]、RT-PCR[6]、荧光原位杂交法[7]等)虽然能够实现细胞组分的灵敏检测,但通常需要对细胞进行固定和裂解等预处理,得到的往往是静态和片段化的信息,难以满足活细胞复杂体系的动态分析需求。为了实现活细胞生物组分的动态检测,分子成像技术应运而生。
荧光成像是基于荧光显微镜的一种分子成像技术,具有直观、无损、高灵敏度、高分辨率等优点。随着荧光成像设备的快速发展,人类探秘自然的视角已经从细胞层面拓展到了亚细胞甚至单分子层面。发展高性能的荧光成像探针,利用荧光显微设备获取靶标物浓度和空间分布信息,已成为化学、生物、医学等领域的热门话题。目前使用较为普遍的荧光成像探针主要有抗体、小分子以及功能核酸等。其中,功能核酸[8]是指具有特殊功能的寡聚核酸分子,由于其具有独特的化学或生物活性,受到了科研工作者的广泛关注。本文主要介绍了近期功能核酸荧光探针在细胞成像中的研究进展。
1. 功能核酸的简介
核酸是由核苷酸单体按照一定顺序连接起来的生物大分子,它是生命体的基本物质,在遗传信息的存储、转录和表达等方面起着重要作用。功能核酸[8]是一类具有特殊化学或生物功能的寡核苷酸分子,包括具有分子识别能力的核酸适体(aptamer)和具有生物催化活性的脱氧核酶(DNAzyme)等。功能核酸的出现突破了人们对传统核酸作为遗传物质的认知,由于其具有高特异性、易于合成和修饰、可设计性高等优点,已成为化学、生物、医学等领域重要的分子工具。
1990年,核酸适体分别由Szostak等和Gold等通过指数富集配体系统进化技术(SELEX)首次筛选得到[9, 10]。在SELEX技术中,首先需要构建一个含大量(1013~1015)随机序列的寡核苷酸文库,然后将该文库与靶标孵育,洗去不能结合靶标或与靶标结合力弱的核酸分子,并通过聚合酶链式反应(PCR)对筛选产物进行扩增。随后,通过多轮筛选富集得到能高特异性、高亲和力结合靶标物的核酸适体,并通过测序获得这些核酸适体的具体序列信息,因此核酸适体又有“化学抗体”之称。核酸适体的筛选靶标物具有普适性,包括金属离子[11]、小分子[12]、氨基酸[13]、肽[14]、蛋白[15]甚至是细胞和组织[16]。此外,核酸适体还具有合成简单、相对分子质量低、化学稳定性高、免疫原性低和易于修饰等优点。这些独特的性质使核酸适体在生化分析领域具有巨大潜力。
随着核酸适体应用的不断发展,核酸适体的筛选方法也在不断更新,Tan等[17, 18]在2006年提出了以活细胞为筛选靶标的Cell-SELEX技术(图 1),该技术基于癌细胞和正常细胞在膜蛋白表达水平上的差异进行核酸适体的筛选。这不仅维持了蛋白组分在细胞生理状态下的构型,还能同时对细胞表面多种生物组分进行筛选,提高了筛选效率。利用该技术,多条能特异性识别靶细胞的核酸适体被筛选出来。例如,他们通过Cell-SELEX技术筛选出能特异性识别胰腺导管癌的核酸适体XQ-2d,该核酸适体能够特异性结合人类胰腺导管癌PL45细胞系(解离常数约为55nmol/L),对临床胰腺导管癌组织样本的检出率高达82.5%,为胰腺导管癌的诊断与治疗提供了具有潜力的分子识别工具[19]。此外,单轮筛选[20]、微流控筛选[21]等筛选技术也在不断发展,有望丰富筛选方法,简化筛选流程,缩短筛选时间。
图 1
随着SELEX技术的不断发展以及对功能核酸源源不断的需求,多种可用于细胞成像的功能核酸被筛选出来,极大推动了其在细胞成像应用中的发展。在此,我们按照成像类别的不同大致将其分为用于细胞外结构域识别的功能核酸和用于细胞内成像的功能核酸(见表 1)。
表 1
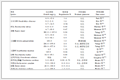 表 1 用于细胞外结构域识别和细胞内成像的功能核酸Table 1. Functional nucleic acids for recognition of extracellular domains and functional nucleic acids for intracellular imaging
表 1 用于细胞外结构域识别和细胞内成像的功能核酸Table 1. Functional nucleic acids for recognition of extracellular domains and functional nucleic acids for intracellular imaging 下载:
导出CSV
下载:
导出CSV
用于细胞识别的功能核酸 用于细胞内成像的功能核酸 靶标 功能核酸 靶标 功能核酸 识别细胞外结构域 神经突触[58] yly12 检测细胞内离子 Zn2+[35] Zn-DNAzyme 细胞隧道纳米管[59] M17A2 Pb2+[66] Pb-DNAzyme 上皮细胞粘附分子[60] SYL3C Na+[67] Na-DNAzyme 核仁素[39] AS1411 检测细胞内小分子 ATP[33] ATPapt 转铁蛋白[55] Tfapt cAMP[68] caDNApt-1 CTLA-4[61] CTLA-4apt 检测细胞内蛋白 cytC[34] cytCapt PDL1[62] PDL1apt NF-kB[57] P028F4 OX40[63] OX40apt P53[57] P53R175H MUC-1[53] 5-TRG2 IFNy[51] IFNγapt CCRF-CEM[17] sgc8 血小板衍生因子[48] PDGFapt PL45[18] XQ-2d 核蛋白[69] Ch4-1 K562[29] T2-KK1B10 凝血酶[70] Thrombinapt Ramos[29] TDO5 细胞生长因子[71] VEGFapt HeLa[64] PP3 HCT116[6] KCHA10 与核酸适体类似,脱氧核酶[22]也是基于SELEX技术筛选得到的。1994年,Breaker和Joyce通过体外筛选方法首次发现了脱氧核酶,它能够在Pb2+存在的情况下催化RNA的切割,这个发现表明单链DNA可以具有类似于核酶和蛋白酶的催化功能。随后的研究发现,除了可用于切割RNA之外,脱氧核酶还可以用来切割DNA[23],连接RNA[24],磷酸化DNA[25]等。一般情况下,脱氧核酶需要结合特定的金属离子或小分子才能启动催化反应,结合适当的信号转导机制,可以对靶标物进行高选择性、高灵敏度检测。相较于常规的蛋白酶和核酶,脱氧核酶具有制备成本低、稳定性高、设计灵活等优点,在金属离子及小分子的生化分析和生物成像领域有广阔的应用前景。
2. 细胞成像
2.1 细胞识别
2.1.1 单价识别
细胞膜蛋白在细胞的生命活动,如细胞增殖和分化、能量转换、信号转导及物质运输等过程中发挥着重要作用。在不同细胞系中,膜蛋白表达具有显著的差异,细胞识别很大程度上可以通过细胞膜蛋白识别得以实现。Cell-SELEX技术是针对细胞膜蛋白差异而进行核酸适体筛选的一种方法,能为细胞识别提供大量有效的分子工具。
Tan等[18]通Cell-SELEX技术筛选得到核酸适体sgc8,并利用该适体实现了对白血病细胞(CCRF-CEM)的特异性识别和荧光成像,为白血病的早期诊断提供了强有力的分子识别工具。为了降低检测背景,提高检测灵敏度,Shi等[26]在此基础上设计了一种激活式探针。将sgc8适体设计为发卡结构,发卡环部序列为识别区域,茎部末端分别修饰上荧光基团和猝灭基团。发卡结构的形成使得荧光基团与猝灭基团彼此靠近,导致荧光猝灭。当其与细胞膜表面的靶标蛋白结合时,发卡型探针发生构象变化,使得荧光基团远离猝灭基团,从而恢复荧光信号,进而实现对目标细胞的特异性识别。他们进一步利用此探针在荷瘤小鼠体内实现了目标癌细胞的高灵敏荧光成像。
2.1.2 多价识别
研究者发现将多个功能核酸修饰在纳米颗粒上或功能核酸自组装等方法能产生多价效应,从而显著提高功能核酸的识别能力。相较于单个功能核酸来说,多价功能核酸具有更高的特异性和亲和性。为了增强免疫细胞与肿瘤细胞之间的相互作用,Shi等[27]在细胞膜表面原位组装多价核酸适体来模拟多价抗体,实验结果表明,相较于单个核酸适体而言,多价核酸适体的亲和性更高,有效提高了免疫细胞对肿瘤细胞的杀伤能力。核酸适体通过折叠成特定的二三级构型,与靶标物特异性结合,通过调节核酸适体的分子识别构型,可实现其对靶标物识别性能的可逆调控,为靶细胞特异性捕获与释放提供了优异的分子工具。为了实现CD8+T细胞的高效捕获,Kacherovsky等[28]将多条核酸适体修饰在磁性纳米颗粒上,通过多价效应最大程度地收集CD8+T细胞,再通过加入核酸适体的互补序列释放和回收所捕获的细胞。
2.1.3 多参数识别
细胞具有高度复杂性和异质性,同时测定多种膜蛋白能有效提高细胞识别的精准性。为了解决单一核酸适体难以准确区分癌细胞类型的难题,进一步实现癌症的智能诊断,Tan等[29]利用核酸适体的分子识别特性,结合DNA链置换反应,发展了一种布尔逻辑回路(图 2(A))。在同一种细胞上实现了多种膜蛋白表达水平的分析。在此基础上,结合适当的治疗试剂(如光敏剂等),可以实现对特定癌细胞的准确诊断和有效治疗[30]。该方法证明了多元核酸适体联用在促进疾病诊断和治疗精准化上的潜力。此外,Tan等[31]还发展了基于多种核酸适体的DNA逻辑回路,结合核酸链式杂交反应(HCR),实现了检测信号的有效放大,提高了检测灵敏度(图 2(B))。
图 2

2.2 细胞内成像
细胞质是细胞进行生命活动的主要场所,为生命活动的正常进行提供能量和原料。对活细胞内重要生物组分的实时成像分析将为细胞生物学的研究提供重要信息。核酸探针具有设计灵活、无需对细胞进行复杂基因操控、无需除去多余探针等优势,是一种颇有前景的细胞内分子成像工具。然而,作为一种带负电荷的亲水性大分子化合物,核酸探针难以跨越细胞膜磷脂双分子层,并且递送至体内的功能核酸大都陷于溶酶体中,这极大限制了其在活细胞内部成像分析中的应用。因此,实现功能核酸有效的递送和溶酶体逃逸,对功能核酸在细胞内部成像分析至关重要。
2.2.1 基于纳米材料的核酸递送方法
纳米材料是指在三维空间中至少有一维处于纳米尺寸(1~100 nm)的新一代材料[32],由于具有组成单元尺度小和光、热性能好等特点,在生物分析和生物成像领域具有十分广阔的前景。研究者发现,纳米材料是性能优良的核酸递送载体,它可以有效负载核酸,并且细胞毒性以及免疫原性较低。此外,有些纳米材料如金纳米颗粒[33]、上转换纳米颗粒、氧化石墨烯纳米片[34]等还能充当良好的猝灭剂,有效降低检测背景,提高检测信噪比。Zheng等[33]通过金硫键将三磷酸腺苷(ATP)特异性响应的核酸适体修饰在金纳米颗粒上,实现核酸适体高效的细胞递送,并对细胞内ATP含量进行了原位检测。He等[35]将近红外响应的双光子脱氧核酶修饰在金纳米颗粒上,实现对细胞内Zn2+的成像分析,该分子探针具有良好的双光子性能和稳定的光性能、较高的组织穿透能力和较低的光毒性。Li等[34]将核酸适体负载在氧化石墨烯纳米片上(图 3(A)),与溶酶体定位剂对比可以看到(图 3(B))纳米片能有效提高功能核酸的溶酶体逃逸效率,以实现对细胞内细胞色素C(Cyt c)动态变化的荧光成像,为细胞凋亡过程研究提供了有效的成像探针。
图 3

通过精确的碱基互补配对,DNA分子可以自组装形成三维纳米结构,如四面体[36]、八面体[37]、三棱柱[38]等。研究发现,DNA分子形成刚性纳米结构后能够显著提高其细胞内吞效率。功能核酸可以通过自组装整合在DNA纳米结构上,随其一起递送至细胞内,对其中的靶标分子进行成像分析,例如,Qiu等[38]将ATP核酸适体劈开并组装在三棱柱上(图 3(C)),可以看到,DNA三棱柱的内吞效率显著高于单链DNA的内吞效率(图 3(D));当DNA三棱柱结合到靶标分子时,组装在上面的两条核酸适体靠近,修饰在核酸适体上的荧光基团也随之靠近,引起高效的荧光能量共振转移(FRET)。该方法可以有效降低假阳性信号,实现细胞内ATP的灵敏检测。
2.2.2 核酸适体靶向
某些核酸适体可以通过与细胞膜表面蛋白特异性结合,在该蛋白介导下进入细胞内。由此可见,功能核酸除了作为检测探针,还可以作为递送探针。基于此,笔者团队利用能够特异性识别癌细胞并可内化的核酸适体作为输送载体,结合分子信标和核酸杂交技术,开发了一种靶细胞自输送的荧光探针(图 4(A)),解决了核酸探针活细胞穿透力差以及常规功能核酸的活细胞递送能力弱的难题[39]。同时,在核酸适体的延伸序列上修饰光响应切割官能团,通过光照精准控制探针的检测性能,从而实现对特定细胞内靶标RNA的高时空分辨成像分析。
图 4

为了实现更精准的细胞靶向递送,Ren等[39]基于核酸适体的智能逻辑运算回路构建了一种DNA双锁的探针设计策略(图 4(C)),通过控制多个参数,提高了细胞识别的特异性,并通过siRNA的高效递送实现了靶标基因的精准沉默,为疾病精准诊疗方法的研究提供了新思路。
2.2.3 模拟荧光蛋白的功能核酸
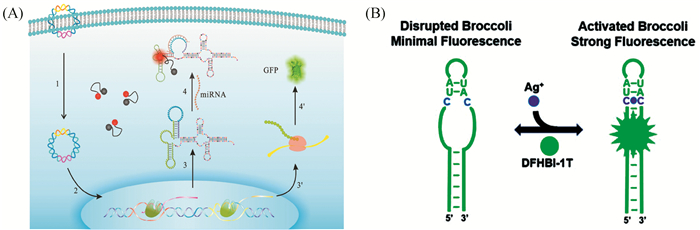
荧光蛋白的出现为蛋白分析提供了一种全新的荧光标记技术,被广泛应用于生物、医学等领域。近年来研究发现了一类核糖核酸适体,其在结合特定的荧光染料分子后能有效恢复该染料分子的荧光。人们利用这种核糖核酸-染料复合物设计了多种“点亮”型荧光探针,包括Spinach[41]、Broccoli[43]、Mango、Core等等。该类“点亮”荧光探针被广泛应用于活细胞中重要靶标分子的荧光成像。例如,Ying等[42](图 5(A))报道了一种新型基因编码核糖核酸探针,用于肿瘤细胞中microRNAs的荧光成像,当探针与靶标microRNA杂交时,发生构型转换,从而与磺基罗丹明荧光染料分子结合,发出高强度荧光。此外,通过核糖核酸适体探针与绿色荧光蛋白(GFP)的共表达,实现了比率成像,这为活细胞内RNA的灵敏成像提供了一种新方法。Yu等[43]报道了一种可用于活细菌内Ag+成像分析的新型基因编码核糖核酸探针(图 5(B))。该探针将胞嘧啶-Ag+-胞嘧啶碱基对引入Broccoli核糖核酸适体中。Ag+-胞嘧啶的结合诱导了Broccoli适体的折叠,并激活了荧光信号,从而实现对细菌中Ag+含量的有效检测。这种合成生物学与分析化学的交叉研究受到越来越广泛的关注,已成为化学生物学领域的一个重要分支。
图 5

2.2.4 细胞内信号放大
细胞内物质的含量普遍较低,一般为10-21~10-15mol[40],一些重要生物组分的含量甚至更低,这给检测带来了极大挑战。功能核酸作为核酸分子,便于结合各种DNA/RNA扩增技术(如滚环扩增反应(RCA)[44]、PCR和恒温循环扩增反应[45]等),使得一个靶标分子可以同时引发多重信号输出,从而实现检测信号的有效放大,以提高检测的灵敏度。为了实现细胞内低丰度miRNA的高灵敏成像,Zhao等[46]报道了一种近红外光激活的HCR(图 6(A))。将修饰有光控基团的分子信标负载在上转换纳米颗粒上,并由此实现分子信标高效的细胞递送。在起始状态下,分子信标被稳定吸附于纳米颗粒表面,处于静默状态。经近红外光照射后,分子信标被激活从而释放出可以发生HCR的粘性末端。当与靶标分子杂交形成双链结构后,分子信标将从颗粒表面脱落下来,并进一步引发HCR反应,实现对靶标分子的信号放大检测。
图 6

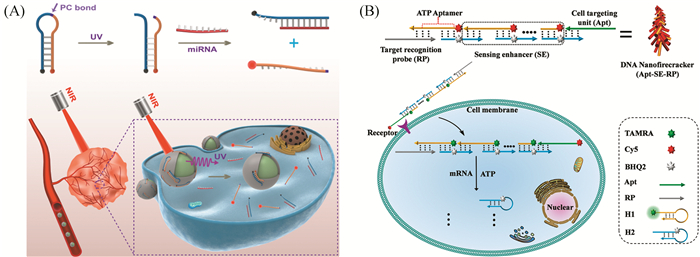
无酶核酸级联反应是一种有效的信号放大策略,然而如何将多种核酸单体运送到细胞内是一大难题。笔者团队受中国鞭炮的启发,设计了一种分子识别激活的可拆卸式纳米鞭炮探针(图 6(B))[47]。该纳米鞭炮探针由三个模块组成,包括细胞靶向模块、分子识别模块以及信号增强模块。纳米探针在靶向模块介导下进入细胞。当分子识别模块与细胞内的靶标mRNA结合后,将从纳米探针上脱落,从而激活了该探针。被激活的纳米探针在细胞内ATP分子的辅助下快速分解,由此产生一对多的信号响应,进而实现对细胞内靶标mRNA的放大成像。
3. 亚细胞器成像
3.1 细胞膜外微环境
细胞膜外微环境(Cellular microenvironment)是指紧密围绕在细胞周围,通过各种物理、化学或生物信号对细胞行为有着直接或间接影响的局部动态环境,在调控细胞生物学功能(包括细胞代谢、繁殖、分化、凋亡及细胞间通讯等)中起着至关重要的作用。实时检测细胞膜外微环境的物质分布,对研究细胞的生命活动具有重要意义。
为了实现对微观环境中重要生理活动的监测,需要将检测探针构建在细胞表面; 由于功能核酸荧光探针具有相对分子质量较小、易被化学修饰等特性,现已被成功应用于细胞膜表面的生物传感。Zhao等[48]将特异性响应血小板生长因子(PDGF)的核酸适体通过共价交联反应固定在间充质干细胞的细胞膜表面,当结合PDGF时,核酸适体构象发生改变,荧光基团相互靠近,发生荧光能量共振转移,产生荧光信号,从而实现了对外加的或相邻细胞分泌的PDGF高时空分辨的定量检测。Tokunaga等[49]通过共价作用将ATP响应的核酸适体固定在脑星形胶质细胞的表面。该探针能够实时监测神经递质ATP的释放;利用该探针还发现ATP的释放信号与钙波在邻近细胞中的传播同步。
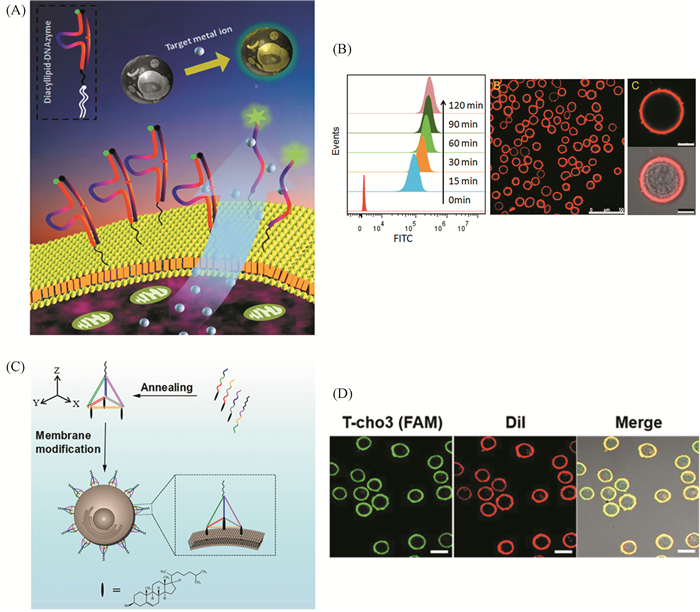
虽然这些荧光探针为细胞膜外微环境的检测提供了有利工具,但它们在细胞膜表面的固定过程均涉及到共价化学反应,可能会影响细胞的生理状态,在一定程度上限制了它们的有效性和实际应用范围。笔者团队在此基础上进行改进,设计合成多种两亲性的脂质链-功能核酸共轭探针,提出一种生物相容性更好、操作更简便以及适用范围更广的细胞膜修饰策略(图 7(A))[50]。该两亲性分子探针通过脂质链与细胞膜磷脂双分子层之间的相似相容作用实现功能核酸在细胞膜表面的高效自组装。通过在细胞膜表面修饰镁离子特异性响应的脱氧核酶探针,对细胞分泌镁离子的生理过程进行实时监测。此外,进一步将此细胞膜表面功能化策略应用于细胞免疫应答过程的研究,结合微流控技术,构建单细胞液滴,在单细胞水平上实现了T细胞应激分泌重要细胞因子生理过程的检测,为细胞免疫应答机制的研究提供了新方法[51]。研究中发现,胆固醇-单链DNA共轭探针在含血清的细胞培养基中容易从细胞膜表面脱落,给其应用研究带来很大干扰。为了解决这个问题,我们在DNA四面体的三个支点上修饰胆固醇分子,成功构建了两亲性三维DNA纳米支架,提高了核酸探针在细胞膜表面的锚定能力(图 7(C))[52]。研究结果表明,相较于胆固醇-单链DNA共轭探针,此两亲性DNA四面体探针在细胞膜表面的锚定能力提高了近100倍,且其靶标识别能力也提高了近2.5倍。此策略有望为多细胞通信网络的研究提供可设计的生物界面传感平台。
图 7

3.2 亚细胞器成像
亚细胞器是细胞中具有特定形态结构和功能的微器官,它们组成了细胞的基本功能结构。随着生命科学的发展以及成像技术的不断更新,科学研究逐渐从细胞层面过渡到亚细胞层面。
Krishnan等[53~55]设计了一系列双链DNA荧光探针,并通过在探针中修饰不同的靶向基团,实现了对溶酶体、高尔基体和质膜等不同亚细胞器中重要生物组分的实时成像。例如,通过引入可特异性结合转铁蛋白的核酸适体,成功将DNA双链荧光探针靶向递送至溶酶体中,实现了溶酶体成熟过程中氯离子的动态检测。实验结果表明,在溶酶体成熟的过程中,其内的氯离子含量逐渐升高; 同时还发现管腔溶酶体氯化物由细胞内氯转运蛋白DmClC-b进行调节,并与溶酶体储存疾病有关[55]。此外,他们通过引入质膜和高尔基内膜的靶向基团,对这两个亚细胞器中NOS3酶的相对含量及活性进行同步成像分析,发现质膜上NOS3酶的活性高于高尔基内膜上,并发现高尔基内膜处的NOS3酶对维持高尔基内膜的形状具有重要意义[53]。由此可见,靶向亚细胞器的分子荧光探针能够提供细胞内生物组分详细的位置信息,从而为细胞生命活动的研究提供更全面信息。
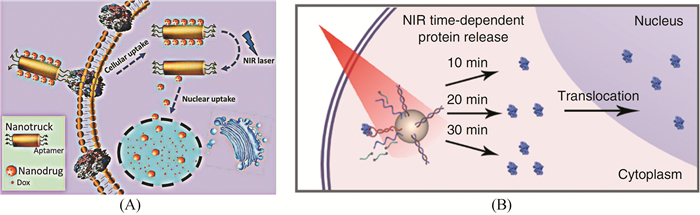
细胞核是细胞遗传与代谢的调控中心,也是大多数抗癌药物分子最终作用的场所。细胞核输送的纳米诊疗探针有望解决由于P-糖蛋白在细胞膜上过表达而引起的癌症多药耐药性问题。然而,细胞膜跨越与细胞核富集对纳米载体的尺寸有着不一样的要求。针对这一问题,笔者等[56]利用核酸适体的分子识别特性,结合DNA自组装技术和生物纳米技术,发展了一种癌细胞靶向和近红外光激活变构的多功能纳米探针,通过金银纳米棒载体的光热效应实现了小尺寸纳米药物的高效解离,基于尺寸效应,可以有效实现纳米探针的细胞核定位和核富集,避免了P-糖蛋白对药物分子的外排作用,大大提高了耐药性癌细胞对药物的敏感性(图 8(A))。同时,通过实时荧光成像示踪为纳米探针的近红外光激活提供了重要指导信息,为癌症示踪介导化疗方法的研究提供了新思路和方法。此外,我们基于核酸适体的近红外光响应纳米平台(图 8(B))设计了一种操控细胞内蛋白亚细胞器定位的策略,利用上转换纳米颗粒作为载体,将能特异性识别细胞调控因子的核酸适体高效递送至细胞质中[57]。核酸适体能特异性捕获Rel A蛋白,并将其截留于细胞质中; 随后,通过近红外光照控制核酸适体的分子识别构型,实现蛋白的定量释放; 被释放的蛋白由此恢复天然的质核穿梭活性。实验结果表明,该纳米光控平台可以有效调节和控制Rel A蛋白在质核之间的穿梭行为,并能影响Rel A相关的下游信号通路。同时该策略还可以用于P53蛋白的线粒体定位。研究表明,野生型P53蛋白可以快速的从细胞质转运到线粒体上,并与Bcl-2家族成员结合,启动内在凋亡途径,突变型的P53蛋白不具备此转移能力,核酸适体P53R175H能与突变型的P53蛋白结合,恢复突变P53蛋白线粒体定位的能力。这种无标签蛋白光调节策略为研究复杂生物事件提供了一个潜力巨大的分子工具。
图 8

4. 结论与展望
功能核酸作为核酸分子具有合成简单、易于修饰、易于结合信号放大策略等特征,还具有相对分子质量低、化学稳定性高、免疫原性低等优点,因此在生化分析、生物成像等领域受到越来越广泛的关注和应用。功能核酸还具有优异的可编程性和可操纵性,使其在生命信息网络研究和操控中有着独特的优势和巨大的应用潜力。
尽管功能核酸探针具有许多优点,目前仍存在一些问题亟需解决。一方面,基于SELEX技术的筛选步骤复杂耗时,目前高性能的功能核酸数量较少,极大地限制了功能核酸的发展;另一方面,功能核酸易被核酸酶降解,在复杂的生物体系中不能长时间稳定存在。目前功能核酸探针在生物医药及生物分析方面的应用才刚刚起步,机遇与挑战并存,我们应该抓住机遇,迎接挑战,努力推动功能核酸在生物分析中的应用。
-
-
[1]
Dean K M, Qin Y, Palmer A E. BBA-Mol. Cell Res., 2012, 1823(9): 1406~1415.
-
[2]
Oddershede L B. Nat. Chem. Biol., 2012, 8(11): 879~886.
-
[3]
Lippincott S J, Snapp E, Kenworthy A. Nat. Rev. Mol. Cell. Biol., 2001, 2(6): 444~456.
-
[4]
Dirks R W, Tanke H J. Biotechniques, 2006, 40(4): 489~496.
-
[5]
Mahmood T, Yang P C. North Am. J. Med. Sci., 2012, 4(9): 429~434.
-
[6]
Deepak S A, Kottapalli K R, Rakwal R, et al. Curr. Genomics, 2007, 8(4): 234~251.
-
[7]
Cui C, Shu W, Li P. Front. Cell Dev. Biol., 2016, 4: 89.
-
[8]
Liu J W, Cao Z H, Lu Y. Chem. Rev., 2009, 109: 1948~1998.
-
[9]
Ellington A D, Szostak J W. Nature, 1990, 346: 818~822.
-
[10]
Tuerk C, Gold L. Science, 1990, 249(4968): 505~510.
-
[11]
Ueyama H, Takagi M, Takenaka S. J. Am. Chem. Soc., 2002, 124(48): 14286~14287.
-
[12]
Baker B R, Lai R Y, Wood M S, et al. J. Am. Chem. Soc., 2006, 128(10): 3138~3139.
-
[13]
Liu L, Stepanian L, Dubins D N, et al. J. Phys. Chem. B, 2018, 122(31): 7647~7653.
-
[14]
Williams K P, Liu X H, Schumacher T N M, et al. PNAS, 1997, 94(21): 11285~11290.
-
[15]
Beatty-Desana J W, Hoggard M J, Cooledge J W. Nature, 1975, 255(5505): 242~243.
-
[16]
Wang C, Zhang M, Yang G, et al. J. Biotechnol., 2003, 102(1): 15~22.
-
[17]
Shangguan D H, Li Y, Tang Z W, et al. PNAS, 2006, 103(32): 11838~11843.
-
[18]
Wu X Q, Liu H L, Han D M, et al. J. Am. Chem. Soc., 2019, 141(27): 10760~10769.
-
[19]
Arnold S, Pampalakis G, Kantiotou K, et al. Biol. Chem., 2012, 393(5): 343~353.
-
[20]
Huang C J, Lin H I, Shiesh S C, et al. Biosens. Bioelectron., 2010, 25(7): 1761~1766.
-
[21]
Tan W H, Donovan M J, Jiang J H. Chem Rev., 2013, 113(4): 2842~2862.
-
[22]
Breaker R R, Joyce G F. Chem. Biol., 1994, 1(4): 223~229.
-
[23]
Santoro S W, Joyce G F, Sakthivel K, et al. J. Am. Chem. Soc., 2000, 122(11): 2433~2439.
-
[24]
Purtha W E, Coppins R L, Smalley M K, et al. J. Am. Chem. Soc., 2005, 127(38): 13124~13125.
-
[25]
Li Y, Breaker R R. PNAS, 1999, 96(6): 2746~2751.
-
[26]
Shi H, He X, Wang K, et al. PNAS, 2011, 108(10): 3900~3905.
-
[27]
Shi P, Wang X, Davis B, et al. Angew. Chem. Int. Ed., 2020, 59(29): 11892~11897.
-
[28]
Kacherovsky N, Cardle I I, Cheng E L, et al. Nat. Biomed. Eng., 2019, 3(10): 783~795.
-
[29]
You M X, Peng L, Shao N, et al. J. Am. Chem. Soc., 2014, 136(4): 1256~1259.
-
[30]
You M X, Zhu G Z, Chen T, et al. J. Am. Chem. Soc., 2015, 137(2): 667~674.
-
[31]
Chang X, Zhang C, Lv C, et al. J. Am. Chem. Soc., 2019, 141(32): 12738~12743.
-
[32]
Biju V. Chem. Soc. Rev., 2014, 43(3): 744~764.
-
[33]
Zheng D, Seferos D S, Giljohann D A, et al. Nano Lett. 2009, 9(9): 3258~3261.
-
[34]
Chen T T, Tian X, Liu C L, et al. J. Am. Chem. Soc., 2015, 137(2): 982~989.
-
[35]
He L, Lu D Q, Liang H, et al. ACS Nano, 2017, 11(4): 4060~4066.
-
[36]
Zhong L, Cai S, Huang Y, et al. Anal. Chem., 2018, 90(20): 12059~12066.
-
[37]
Zheng X F, Peng R Z, Jiang X, et al. Anal. Chem., 2017, 89(20): 10941~10947.
-
[38]
Qiu L P, Wu C C, You M X, et al. J. Am. Chem. Soc., 2013, 135(35): 12952~12955.
-
[39]
Ren K, Liu Y, Wu J, et al. Nat. Commun., 2016, 7: 13580.
-
[40]
De Souza N. Nat. Methods, 2012, 7: 35.
-
[41]
Jeremy S P, Karen W, Samie R. Science, 2011, 333(6042): 642~646.
-
[42]
Ying Z M, Wu Z, Tu B, et al. J. Am. Chem. Soc., 2017, 139(29): 9779~9782.
-
[43]
Yu Q, Shi J, Mudiyanselage A P K K K, et al. Chem. Commun., 2019, 55(5): 707~710.
-
[44]
Deng R, Tang L, Tian Q, et al. Angew. Chem. Int. Ed., 2014, 53(9): 2389~2393.
-
[45]
He L, Lu D Q, Liang H, et al. J. Am. Chem. Soc., 2018, 140(1): 258~263.
-
[46]
Zhao J, Chu H, Zhao Y, et al. J. Am. Chem. Soc., 2019, 141(17): 7056~7062.
-
[47]
Xing X J, Li J, Qiu L P, et al. Chem. Commun., 2020, 56(21): 3131~3134.
-
[48]
Zhao W, Schafer S, Choi J, et al. Nat. Nanotechnol., 2011, 6(8): 524~531.
-
[49]
Tokunaga T, Namiki S, Yamada K, et al. J. Am. Chem. Soc., 2012, 134(23): 9561~9564.
-
[50]
Qiu L P, Zhang T, Jiang J H, et al. J. Am. Chem. Soc., 2014, 136(38): 13090~13093.
-
[51]
Qiu L P, Wimmers F, Weiden J, et al. Chem. Commun., 2017, 53(57): 8066~8069.
-
[52]
Li J, Xun K Y, Pei K, et al. J. Am. Chem. Soc., 2019, 141(45): 18013~18020.
-
[53]
Jani M S, Zou J, Veetil A T, et al. Nat. Chem. Biol., 2020, (16): 660~666.
-
[54]
Modi S, Goswami D, Gupta G D, et al. Nat. Nanotechnol., 2009, (4): 325~330.
-
[55]
Saha S, Prakash V, Halder S, et al. Nat. Nanotechnol., 2015, (10): 645~651.
-
[56]
Qiu L P, Chen T, Ocsoy I, et al. Nano Lett., 2015, 15(1): 457~463.
-
[57]
Xie S T, Du Y L, Zhang Y, et al. Nat. Commun., 2020, 11(1): 1347.
-
[58]
Wang L, Bing T, Liu Y, et al. J. Am. Chem. Soc., 2018, 140(51): 18066~18073.
-
[59]
Zhang N, Bing T, Shen L, et al. Angew. Chem. Int. Ed., 2016, 55(12): 3914~3918.
-
[60]
Gutsaeva D, Parkerson J B, Yerigenahally S, et al. Blood, 2011, 117(2): 727~735.
-
[61]
Santulli-Marotto S, Nair S K, Rusconi C, et al. Cancer Res., 2003, 63(21): 7483~7489.
-
[62]
Prodeus A, Abdul-Wahid A, Fischer N W, et al. Mol. Ther-Nucl Acids, 2015, 4: e237.
-
[63]
Dollins C M, Nair S, Boczkowski D, et al. Chem. Biol., 2008, 15(7): 675~682.
-
[64]
Parekh P, Tang Z, Turner P C, et al. Anal. Chem., 2010, 82(20): 8642~8649.
-
[65]
Sefah K, Meng L, Lopez-Colon D, et al. PLoS One, 2010, 5(12): e14269~e14279.
-
[66]
Cui L, Peng R, Fu T, et al. Anal. Chem., 2016, 88(3): 1850~1855.
-
[67]
Chen K, Huang Q, Fu T, et al. Anal. Chem., 2020, 92(11) 7404~7408.
-
[68]
Sharma S, Zajac M, Krishnan Y. ChemBioChem, 2020, 21(1-2): 157~162.
-
[69]
Shen L, Bing T, Zhang N, et al. ACS Sensors, 2019, 4(6): 1612~1618.
-
[70]
Schultze P, Macaya R F, Feigon J. J. Mol. Biol., 1994, 235(5): 1532~1547.
-
[71]
Ng W M, Shima D T, Calias P, et al. Nat. Rev. Drug Discov., 2006, 5(2): 123~132.
-
[1]
-
表 1 用于细胞外结构域识别和细胞内成像的功能核酸
Table 1. Functional nucleic acids for recognition of extracellular domains and functional nucleic acids for intracellular imaging
用于细胞识别的功能核酸 用于细胞内成像的功能核酸 靶标 功能核酸 靶标 功能核酸 识别细胞外结构域 神经突触[58] yly12 检测细胞内离子 Zn2+[35] Zn-DNAzyme 细胞隧道纳米管[59] M17A2 Pb2+[66] Pb-DNAzyme 上皮细胞粘附分子[60] SYL3C Na+[67] Na-DNAzyme 核仁素[39] AS1411 检测细胞内小分子 ATP[33] ATPapt 转铁蛋白[55] Tfapt cAMP[68] caDNApt-1 CTLA-4[61] CTLA-4apt 检测细胞内蛋白 cytC[34] cytCapt PDL1[62] PDL1apt NF-kB[57] P028F4 OX40[63] OX40apt P53[57] P53R175H MUC-1[53] 5-TRG2 IFNy[51] IFNγapt CCRF-CEM[17] sgc8 血小板衍生因子[48] PDGFapt PL45[18] XQ-2d 核蛋白[69] Ch4-1 K562[29] T2-KK1B10 凝血酶[70] Thrombinapt Ramos[29] TDO5 细胞生长因子[71] VEGFapt HeLa[64] PP3 HCT116[6] KCHA10  下载: 导出CSV
下载: 导出CSV
-












 扫一扫看文章
扫一扫看文章
计量
- PDF下载量: 30
- 文章访问数: 2334
- HTML全文浏览量: 709
