
Figure 1.
SEM images of TiNFs (A, B) and TEM images of TiNTs (C, D).

The development of nanotechnologies leads to the wide use of nanoparticles (NPs) in modern society, and therefore direct contact with engineering NPs is highly possible in daily life [1]. In particular, NPs without acute toxic effects have been attracted extensive research interests, especially in biomedical fields [2-4]. Among the engineering NPs, TiO2 NPs have been widely used in many areas ranging from photocatalysis [5] to food industry [6] to biomedicine [7]. Although TiO2 NPs were originally considered as biologically inert nanomaterials (NMs), extensive studies still revealed that exposure to TiO2 NPs could damage cell models in vitro and animal models in vivo, which highlighted a need to carefully evaluate the threat of TiO2 NP exposure to human beings [8, 9]. Likewise TiO2 NPs, titanate-based NMs, such as titanate nanofibers (TiNFs) [10], titanate nanotubes (TiNTs) [11], titanate nanoflaks [12] and titanate nanocomplexes [13] have been widely used in similar disciplines. Meanwhile, TiNFs and TiNTs have been proposed as biocompatible materials for biomedical uses [14, 15]. As such, it is expected that human beings will be increasingly exposed to titanate-based NMs. But at present, the toxicity of TiNFs and TiNTs was less systemically investigated as compared with TiO2 NPs.
Exposure to NMs will likely lead to the entering of NMs into circulation, and therefore it is necessary to investigate the toxicity of NMs to cells lining blood vessels, in particular endothelial cells (ECs), the surface cells covering the lumen of blood vessels [16]. Indeed, the vascular toxicity of TiO2 NPs has been extensively studied before, but the mechanisms are not fully known yet [17, 18]. However, the vascular toxicity and the mechanisms of TiNFs and TiNTs were less investigated yet. Kruppel-like factors (KLF) are a family of transcription factors (TF) crucial in the regulation of vascular biology, and therefore investigating the changes of KLF can help the understanding of adverse vascular effects of pathological and environmental stimuli [19, 20]. Interestingly, we recently reported that different types of NMs altered KLF in both human ECs [21-23] and vascular smooth muscle cells [24, 25]. However, the downstream events were not clear. Since TF including KLF could regulate many downstream events, it may be necessary to use systems biology methods (for example omics-techniques and bioinformatics) to investigate KLF-mediated effects due to NM exposure [26].
In this study, we exposed an endothelial monolayer model to TiNFs and TiNTs, and performed transcriptomic analysis to investigate the influence of NMs on KLF and KLF-regulated downstream events and signaling pathways. The TiNFs (product code XFJ47) and TiNTs (code XFJ46) used in this study were purchased from Nanjing XFNANO Materials Tech Co., Ltd. According to the data provided by the supplier, the diameter, length and surface area of TiNFs are 40−500 nm, 5−20 μm and 20-30 m2/g, respectively. For TiNTs, the diameter, length and surface area are 4−10 nm, 100−500 nm and 200-300 m2/g, respectively. The scanning electron microscope (SEM) images shown in Figs. 1A and B suggest that TiNFs are relatively long and wide fibers, whereas transmission electron microscope (TEM) images shown in Figs. 1C and D suggest that TiNTs are short and small tubes.
In this study, we developed endothelial monolayer model by using cell culture inserts to better mimic the endothelium in vivo [27]. To this end, human umbilical vein endothelial cells (HUVECs) were cultivated onto cell culture inserts for long term (totally 5 days) to develop polarized endothelial monolayer and then exposed to TiNFs and TiNTs for another 24 h (Fig. S1 in Supporting information). The results from cell counting kit-8 (CCK-8) assay indicated that incubation with various concentrations of TiNFs or TiNTs did not significantly decrease the cellular viability of endothelial monolayer (Fig. S2 in Supporting information). Both TiNFs (Fig. S3A in Supporting information) and TiNTs (Fig. S3B in Supporting information) dose-dependently increased cellular Ti elemental concentrations, but TiNFs led to much higher levels of Ti elemental concentrations in endothelial monolayer compared with TiNTs. Previously we found that TiNTs did not significantly induce cytotoxicity to 2 D HUVEC monoculture [28]. Meanwhile, we also reported that TiO2 NPs did not induce cytotoxic effects in HUVECs [29] or vascular smooth muscle cells [25]. The results obtained in this study are consistent with our previous studies and could indicate the relatively high biocompatibility of Ti-based NMs to vascular cells. Another reason for lack of cytotoxicity could be due to that we used endothelial monolayer model developed on cell culture inserts, and this model better resembles tissues and is generally more resistant to NM exposure [27, 30].
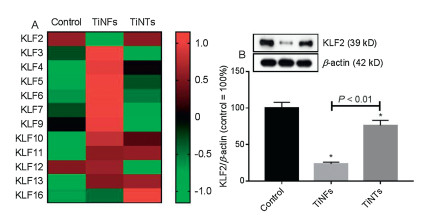
We then did transcriptomics to investigate the systemic effects of TiNFs and TiNTs. The results from transcriptomics indicated that TiNFs significantly up-regulated 235 genes and down-regulated 177 genes, with 58, 413 genes not significantly altered (Fig. S4A in Supporting information). TiNTs significantly up-regulated 174 genes and down-regulated 444 genes, with 58, 207 genes not significantly altered (Fig. S4B in Supporting information). We then focused on the changes of KLF, because KLF are important in regulation vascular biology [19, 20]. Meanwhile, we showed before even NMs with no obvious cytotoxicity could influence KLF in vascular cells. For example, PEG-functionalized Au nanorods were not cytotoxic but decreased KLF2 leading to increased soluble monocyte chemotactic protein 1 (sMCP-1) release in HUVECs [21]. Similarly, TiNTs and TiO2 NPs were not cytotoxic but reduced KLF2-KLF4-eNOS pathways in HUVECs [28]. In another study, we showed that halloysite nanotubes only induced modest cytotoxic effects but reduced KLF2 proteins leading to endothelial activation both in HUVECs and aortas of mice [23]. However, in the previous studies we used traditional toxicological methods, but those methods may not systemically investigate the influence of NMs on KLF and the downstream events. Here by analyzing transcriptomic data, we found that totally 12 KLF genes were expressed in at least one group (genes with FPKM value larger than 1 were considered as expressed; Fig. 2A). Among them, KLF4 and KLF5 were not expressed in control group but up-regulated following TiNT or TiNF exposure. The other 10 KLF genes were expressed in all groups. Most of the KLF genes, such as KLF3, KLF6 and KLF7, were up-regulated by TiNFs but not TiNTs. KLF11 was significantly up-regulated by both TiNFs (P= 1.5×10−5) and TiNTs (P= 3.6×10−5). KLF16 was only significantly up-regulated by TiNTs (P= 3.9×10-4) but not TiNFs (P > 0.05). KLF2 was significantly down-regulated by TiNFs (P= 9.36×10-4) but not TiNTs (P= 0.74). To confirm the changes of KLF2, Western blot was used to measure KLF2 proteins. As shown in Fig. 2B, KLF2 proteins were significantly down-regulated by both TiNFs and TiNTs (P < 0.01), but TiNFs led to significantly lower levels of KLF2 proteins compared with TiNTs (P < 0.01). In this study we focused on the changes of KLF2 because KLF2 is a well-documented TF that could elicit protective effects to ECs and blood vessels [19, 20]. Hence, a significant decrease of KLF2 by TiNFs is expected to mediate adverse vascular effects. Compared with our previous report [28], it appeared that the influence of TiNTs on KLF2 was less effective. This may be due to that we used monolayer model rather than 2 D monoculture, and the monolayer model is more resistant to NM exposure [27].
Then, we did bioinformatics to investigate the possible downstream events due to decreased KLF2 levels. Ingenuity pathway analysis (IPA) database suggested that 196 genes were downstream genes of KLF2. Among them, totally 110 genes were expressed in all groups (FPKM > 1), and 3 genes (F3, vascular cell adhesion molecule 1 (VCAM1) and parathyroid hormone like hormone (PTHLH)) were only expressed in TiNF-exposed but not control or TiNT-exposed groups. Among the KLF2-regulated genes, 10 genes were significantly altered by at least one type of NMs (shown in Fig. S5 in Supporting information). Typically, most of these genes, such as CCL2 (C-C motif chemokine ligand 2), CXCL2 (C-X-C motif chemokine ligand 2) and CXCL8, are related with inflammatory responses and were only significantly up-regulated by TiNFs but not TiNTs. CLDN5 (claudin 5) was significantly up-regulated by both TiNFs (P= 0.028) and TiNTs (P= 9.8×10−13), whereas TFRC (transferrin receptor) was significantly down-regulated by both TiNFs (P= 6.2×10-5) and TiNTs (P= 5.9×10-6).
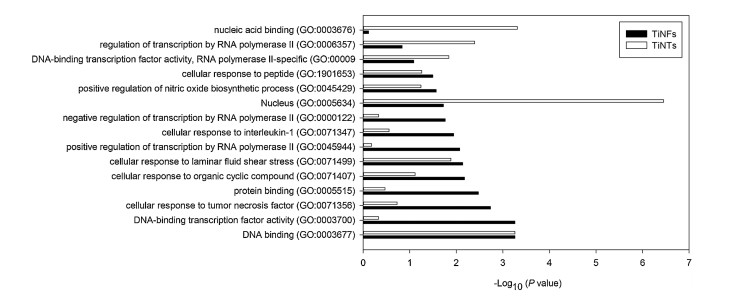
Among KLF2-related GO terms, totally 15 GO terms were significantly affected by at least one type of NMs (Fig. 3). Most of the GO terms, such as positive regulation of nitric oxide biosynthetic process (GO: 0045429), cellular response to tumor necrosis factor (GO: 0071356) and cellular response to interleukin-1 (GO: 0071347), were only significantly altered by TiNTs (P < 0.05) but not TiNTs (P > 0.05). Some GO terms, such as DNA binding (GO: 0003677), cellular response to laminar fluid shear stress (GO: 0071499) and nucleus (GO: 0005634), were significantly altered by both TiNFs and TiNTs (P < 0.05). The GO terms DNA-binding transcription factor activity, RNA polymerase Ⅱ-specific (GO: 0000981), regulation of transcription by RNA polymerase Ⅱ (GO: 0006357) and nucleic acid binding (GO: 0003676) were only significantly altered by TiNTs (P < 0.05) but not TiNFs (P > 0.05).
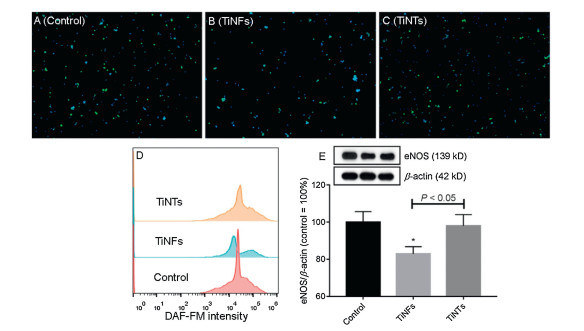
KLF2 is involved in three KEGG pathways, namely FoxO signaling pathway (ko04068), apelin signaling pathway (ko04371) and fluid shear stress and atherosclerosis (ko05418). Among these KEGG pathways, only fluid shear stress and atherosclerosis (ko05418) pathway was significantly altered by TiNFs (P= 0.03) but not TiNTs (P= 0.25). The other two KEGG pathways were not significantly altered by TiNFs or TiNTs (P > 0.05). The KEGG pathway map of fluid shear stress and atherosclerosis (ko05418) indicates that KLF2 can positively regulate eNOS to mediate beneficial effects to ECs, such as anti-inflammatory, anti-thrombotic, anti-oxidant and vasodilatory effects (Fig. S6 in Supporting information). The above results led us to speculate that TiNFs can decrease KLF2-eNOS leading to insufficient NO production and increased inflammatory gene expression. To confirm this, the levels of NO and eNOS were measured. As shown in Figs. 4A-C, at least some cells from all groups could be stained by NO probe DAF-FM with green fluorescence. Quantitative analysis showed that NO was decreased slightly by TiNTs and more markedly by TiNFs (Fig. 4D). The protein levels of eNOS were significantly decreased by TiNFs (P < 0.05) but not TiNTs (P > 0.05), and TiNFs led to significantly lower levels of eNOS compared with TiNTs (P < 0.05;Fig. 4E). Previous studies by using animal models showed that exposure to TiO2 NPs induced atherosclerotic changes due to insufficient NO production leading to endothelial dysfunction [31-33]. However, the data obtained by using in vitro models appeared to be inconsistent, as decreased [34], increased [35] or even unaltered [36] eNOS levels and NO production had been observed in TiO2 NP-exposed ECs. Hereby, by using endothelial monolayer model which better mimics in vivo endothelium [27], we showed that TiNFs decreased NO production due to the down-regulation of KLF2. TiNFs showed larger impact on KLF2-eNOS pathway compared with TiNTs, which may be due to that TiNFs more effectively led to Ti element accumulation compared with TiNTs (Figs. S3A and B).
In summary, this study showed that exposure to TiNFs or TiNTs did not cytotoxicity to endothelial monolayer model despite substantial NM internalization. However, combining transcriptomics and traditional toxicological analysis, we revealed that TiNFs and more modestly TiNTs decreased KLF2 leading to decreased eNOS proteins and NO production. At present, many NPs without acute toxicity, including TiNFs and TiNTs, are being investigated for various biomedical applications [15, 37-39]. Our data may provide novel understanding about the adverse health effects of non-cytotoxic NMs to ECs.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was financially supported by Hunan Innovative Province Construction Special Major Landmark Innovation Demonstration Project (No. 2019XK2303) and Xiangtan Science and Technology Project (No. ZD-ZD20191007).
Supplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/j.cclet.2020.10.044.
M.E. Vance, T. Kuiken, E.P. Vejerano, et al., Beilstein J. Nanotechnol. 6 (2015) 1769-1780. doi: 10.3762/bjnano.6.181
H. Huang, L. Mao, Z. Li, et al., J. Bioresour. Bioprod. 4 (2019) 231-241.
P. Miao, B. Wang, Z. Yu, J. Zhao, Y. Tang, Biosens. Bioelectron. 63 (2015) 365-370. doi: 10.1016/j.bios.2014.07.075
W. Yu, M. Shevtsov, X. Chen, H. Gao, Chin. Chem. Lett. 31 (2020) 1366-1374. doi: 10.1016/j.cclet.2020.02.036
C. Dong, J. Ji, Z. Yang, et al., Chin. Chem. Lett. 30 (2019) 853-862. doi: 10.1016/j.cclet.2019.03.020
J. Musial, R. Krakowiak, D.T. Mlynarczyk, T. Goslinski, B.J. Stanisz, Nanomater. (Basel) 10 (2020) 1110. doi: 10.3390/nano10061110
H. Huang, W. Feng, Y. Chen, J. Shi, Nano Today 35 (2020) 100972. doi: 10.1016/j.nantod.2020.100972
Z. Luo, Z. Li, Z. Xie, et al., Small 16 (2020) e2002019. doi: 10.1002/smll.202002019
C. Bai, M. Tang, J. Appl, Toxicol. 40 (2020) 37-63.
Q. Chen, L. Chen, J. Qi, et al., Chin. Chem. Lett. 30 (2019) 1214-1218. doi: 10.1016/j.cclet.2019.03.002
T. Zheng, T. Wang, R. Ma, et al., Sci. Total Environ. 650 (2019) 1412-1418. doi: 10.1016/j.scitotenv.2018.09.152
W. Liu, W. Zhang, M. Liu, et al., Chin. Chem. Lett. 30 (2019) 2177-2180. doi: 10.1016/j.cclet.2019.07.050
X. Zhang, Y. Guo, Y. Li, Y. Liu, S. Dong, Chin. Chem. Lett. 30 (2019) 502-504. doi: 10.1016/j.cclet.2018.07.007
A. Ozkizilcik, R. Williams, Z.R. Tian, et al., Methods Mol. Biol. 1727 (2018) 433-442.
Y. Ranjous, G.J. Regdon, K. Pintye-Hódi, T. Sovány, Drug Discov. Today 24 (2019) 1704-1709. doi: 10.1016/j.drudis.2019.05.019
Y. Cao, Y. Gong, L. Liu, et al., J. Appl. Toxicol. 37 (2017) 1359-1369. doi: 10.1002/jat.3470
Y. Cao, Y. Gong, W. Liao, et al., BioMetals. 31 (2018) 457-476. doi: 10.1007/s10534-018-0113-7
P. Møller, L. Mikkelsen, L.K. Vesterdal, et al., Crit. Rev. Toxicol. 41 (2011) 339-368. doi: 10.3109/10408444.2010.533152
Y. Fan, H. Lu, W. Liang, W. Hu, J. Zhang, Y.E. Chen, J. Mol. Cell Biol. 9 (2017) 352-363. doi: 10.1093/jmcb/mjx037
D.R. Sweet, L. Fan, P.N. Hsieh, M.K. Jain, Front. Cardiovasc. Med. 5 (2018) 6. doi: 10.3389/fcvm.2018.00006
X. Li, Y. Tang, C. Chen, D. Qiu, Y. Cao, Toxicol. Appl. Pharmacol. 382 (2019) 114758. doi: 10.1016/j.taap.2019.114758
Z. Li, T. Liu, J. Long, et al., Chin. Chem. Lett. 30 (2019) 582-586. doi: 10.1016/j.cclet.2018.12.011
B. Wu, M. Jiang, X. Liu, et al., Nanotoxicology 14 (2020) 1017-1038. doi: 10.1080/17435390.2020.1780642
H. Yang, J. Li, C. Yang, H. Liu, Y. Cao, Toxicol. Appl. Pharmacol. 374 (2019) 11-19. doi: 10.1016/j.taap.2019.04.022
M. Wang, Q. Yang, J. Long, et al., Int. J. Nanomedicine 13 (2018) 8037-8049. doi: 10.2147/IJN.S188175
R. Chen, J. Qiao, R. Bai, Y. Zhao, C. Chen, Anal. Bioanal. Chem. 410 (2018) 6051-6066. doi: 10.1007/s00216-018-0940-y
Y. Cao, S. Li, J. Chen, Toxicol. Mech. Methods 31 (2021) 1-17. doi: 10.1080/15376516.2020.1828521
S. Li, H. Liu, Z. Zhou, Y. Cao, Toxicol. In Vitro 62 (2020) 104689. doi: 10.1016/j.tiv.2019.104689
Y. Gu, S. Chen, G. Chen, et al., Toxicol. Mech. Methods 27 (2017) 191-200. doi: 10.1080/15376516.2016.1273429
D. Zink, J.K.C. Chuah, J.Y. Ying, Trends Mol. Med. 26 (2020) 570-582. doi: 10.1016/j.molmed.2020.01.008
L. Mikkelsen, M. Sheykhzade, K.A. Jensen, et al., Part. Fibre Toxicol. 8 (2011) 32. doi: 10.1186/1743-8977-8-32
T. Chen, J. Hu, C. Chen, et al., J. Nanosci. Nanotechnol. 13 (2013) 3214-3222. doi: 10.1166/jnn.2013.7147
X. Yu, X. Zhao, Y. Ze, et al., J. Hazard. Mater. 280 (2014) 364-371. doi: 10.1016/j.jhazmat.2014.08.015
N. Bayat, V.R. Lopes, J. Schölermann, L.D. Jensen, S. Cristobal, Biomaterials 63 (2015) 1-13. doi: 10.1016/j.biomaterials.2015.05.044
M.D.P. Ramos-Godínez, B.E. González-Gómez, A. Montiel-Dávalos, R. LópezMarure, E. Alfaro-Moreno, Toxicol. In Vitro 27 (2013) 774-781. doi: 10.1016/j.tiv.2012.12.010
Z. Gholinejad, M.H. Khadem Ansari, Y. Rasmi, J. Trace Elem. Med. Biol. 54 (2019) 27-35. doi: 10.1016/j.jtemb.2019.03.008
Z. Chang, F. Liu, L. Wang, et al., Chin. Chem. Lett. 30 (2019) 1856-1882. doi: 10.1016/j.cclet.2019.08.034
R. Miraftab, H. Xiao, J. Bioresour. Bioprod. 4 (2019) 200-201.
F. An, N. Chen, W.J. Conlon, et al., Int. J. Biol. Macromol. 153 (2020) 100-106. doi: 10.1016/j.ijbiomac.2020.02.253

Figure 2 The changes of KLF family. Endothelial monolayer was incubated with cell culture medium (control), 64 μg/mL TiNFs or TiNTs for 24 h. (A) The heat-map was made based on average FPKM values of KLF genes. These KLF genes were expressed in at least one group (FPKM values larger than 1 were considered as expressed). (B) Western blot was used to measure the protein levels of KLF2. *P < 0.01, compared with control.

Figure 3 The P values of GO terms (-Log10 transformed). These GO terms are related with KLF2 and were significantly altered by at least one type of NMs.

Figure 4 The changes of NO-eNOS. Endothelial monolayer was incubated with cell culture medium (control), 64 μg/mL TiNFs or TiNTs for 24 h. The intracellular NO and nuclei were stained with green fluorescence by DAF-FM and blue fluorescence by Hoechst 33342, respectively. (A-C) The representative fluorescence images of control (A), TiNF- (B) and TiNT-exposed (C) exposed cells. (D) The changes of DAF-FM intensity. (E) The protein levels of eNOS were measured by Western blot. *P < 0.05, compared with control.

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 DownLoad:
DownLoad:

 下载:
下载: