State Key Laboratory of Advanced Medical Materials and Devices, Tianjin Key Laboratory of Biomedical Materials, Key Laboratory of Biomaterials and Nanotechnology for Cancer Immunotherapy, Institute of Biomedical Engineering, Chinese Academy of Medical Sciences and Peking Union Medical College, Tianjin 300192, China
c.
Guangdong Laboratory of Artificial Intelligent and Digital Economy (SZ), Shenzhen 518107, China
Received Date:
06 July 2024 Accepted Date:
03 November 2024 Revised Date:
30 October 2024 Available Online:
15 August 2025
Abstract:
Neuronal mitochondrial damage is the primary characteristic of Alzheimer's disease (AD); as mitochondrial microRNAs (miRNAs) are crucial for maintaining mitochondrial function, developing a detection system for mitochondrial miRNAs and applying it for AD treatment is of great significance. Herein, we report CeO2-DNA-RNA hybrid chain (DRP)/mitochondrial aptamers (MA) nanoclusters, formed using specially modified 5 nm sized cerium dioxide (CeO2) nanoparticles, to achieve AD diagnosis and treatment by targeting mitochondrial miRNA-204. First, nanomaterials with unique reactive oxygen species-scavenging functions could easily enter the central nervous system, and surface modification with mitochondrial aptamers facilitated successful mitochondrial targeting. Furthermore, surface modification of nanomaterials with DNA-RNA hybrid biological detection probes was used to detect miRNA-204 in AD and simultaneously perform gene-silencing therapy. In 3×Tg-AD model mice, CeO2-DRP/MA aggregated into neuronal mitochondria, silenced mitochondrial miRNA-204, and restored the damaged mitochondria for AD treatment. The promising diagnostic and therapeutic functions of CeO2-DRP/MA demonstrate its better performance as a diagnostic and therapeutic system targeting mitochondrial miRNA.
Alzheimer's disease (AD) is a common neurological illness that causes cognitive impairment in older individuals [1–6]. AD affects approximately 50 million older individuals worldwide, with a significant prevalence among individuals aged > 60 years. Despite four hypotheses on AD pathogenesis being proposed, the specific pathogenetic mechanism of AD remains uncertain [7–12]. Drug development for AD treatment is currently ongoing worldwide. However, because AD affects the nervous system and has an extremely complex pathogenesis, significant challenges remain in the development of AD drugs. Consequently, advanced approaches are needed for the early diagnosis and treatment of AD.
Cellular autophagy plays a critical role in AD pathogenesis. This process precedes apoptosis and initiates apoptotic pathways when cells are unable to avoid death through the autophagic responses [13–18]. Early and late stage autophagic vacuoles (AVs) in AD brain tissue are rich in light chain 3 Ⅱ (LC3-Ⅱ), and the level of LC3-Ⅱ is significantly elevated [19]. Moreover, mitochondrial autophagy is closely related to the excessive production of reactive oxygen species in cells. Mitochondria miRNA-204 is highly expressed in AD and triggers cellular autophagy by controlling mitochondrial energy output, leading to mitochondrial dysfunction [20–24]. Transient receptor potential mucolipin 1 (TRPML1), a member of the miRNA-204 family, has been found to be a crucial gene in regulating AD pathogenesis. miRNA-204 affects the occurrence of cellular autophagy by regulating the signal transducer and activator of transcription 3 (STAT3) pathway [25–27]. Therefore, detection and regulation of mitochondrial miRNA-204 is particularly important for AD diagnosis and treatment.
Cerium dioxide (CeO2) exists excellent thermal and chemical stability, as well as electronic interactions with other components, which is commonly used as a carrier material or additive to enhance the stability and activity of nanomaterials [28–31]. The unique Ce4+/Ce3+ redox cycle in CeO2 nanomaterials endows them with the activity of several natural oxidoreductases [32–35]. Therefore, CeO2 nanomaterials can eliminate reactive oxygen species and prevent cellular damage in autophagy, which is usually accompanied by an abnormal increase in reactive oxygen species. Overall, these advantages make CeO2 as an effective nanoplatform for drug delivery.
In this study, CeO2-DRP/MA nanomaterials were composed of aminated 5 nm sized CeO2 nanoparticles as the core, which were covalently linked with mitochondrial aptamers (MA) and DNA-RNA hybrid chains (DRP) to detect mitochondrial miRNA-204 on their surface (Fig. 1a). DNA chain contains antisense oligonucleotides (ASO) functional chain, which can silence the target miRNA-204. By binding to its complementary target mRNA and using the mechanism of complementary pairing editing, which is facilitated by ribonuclease H1, ASO can stifle the expression of its target gene. After the DNA-RNA hybrid chains in the nanomaterials specifically recognized mitochondrial miRNA-204, RNA chains with quenched groups were competitively dropped, while the carboxyfluorescein (FAM) at the end of the DNA chain in the hybrid chain underwent fluorescence recovery for detection. Meanwhile, owing to the formation of a new double-stranded DNA-RNA hybrid chain containing the DNA strand and the mitochondrial miRNA-204, mitochondrial miRNA-204 in the double-stranded structure was cleaved by endogenous RNA enzymes. Therefore, mitochondrial miRNA-204 was silenced, and the mitochondria-related autophagy was regulated. This caused damaged cells to move away from autophagy and return to their normal state, ultimately improving the symptoms of AD.
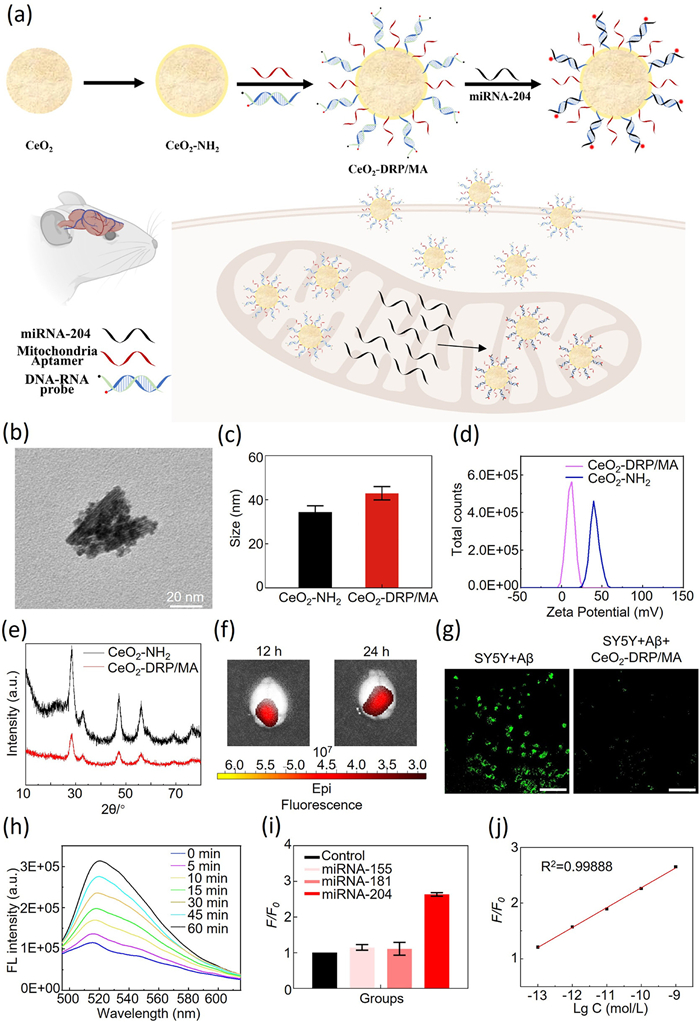
Figure 1
Figure 1.
(a) Synthetic route of CeO2-DRP/MA and schematic diagram of CeO2-DRP/MA use in miRNA-204 biomarker detection and AD therapy. (b) HR-TEM image of CeO2-DRP/MA. Scale bar: 20 nm. (c) Size analysis of CeO2—NH2 and CeO2-DRP/MA. (d) Zeta potential analysis of CeO2—NH2 and CeO2-DRP/MA. (e) X-ray diffraction patterns of CeO2—NH2 and CeO2-DRP/MA. (f) Fluorescence imaging of 3×Tg-AD mouse brains at various time points after CeO2-DRP/MA injection. (g) Active oxygen removal ability based on CLSM images of different group cells incubated with CeO2-DRP/MA for 24 h (λex = 495 nm, λem = 529 nm). Scale bar: 100 µm. (h) Fluorescence intensities of CeO2-DRP/MA in the presence of miRNA-204 in solution at different time intervals. (i) The detection specificity of CeO2-DRP/MA for mitochondrial miRNA-204. (j) The detection sensitivity of CeO2-DRP/MA for mitochondrial miRNA-204. Data in c, i are presented as mean ± standard deviation (SD) (n = 3).
CeO2-DRP/MA nanomaterials were composed of 5 nm sized CeO2 nanoparticles linked with MA and DRP on their surface. High resolution transmission electron microscope (HR-TEM) was used to characterize the morphology and size of CeO2-DRP/MA nanomaterials. Fig. 1b showed a CeO2-DRP/MA nanocluster structure composed of 5 nm circular CeO2 nanoparticles, confirming that the average size of the prepared nanocluster material was approximately 43 nm. The particle size and zeta potential of the nanomaterials was further confirmed using dynamic light scattering (DLS). As shown in Fig. 1c, the particle size of CeO2—NH2 was 37.18 nm, and after surface modification with MA and DRP, the particle size of the nanomaterial increased to 43.59 nm. Thus, the connection between MA and DRP increased the particle size of CeO2—NH2, indicating successful assembly of the nanomaterial. In Fig. 1d, the potential of CeO2—NH2 was approximately 48.88 mV while that of CeO2-DRP/MA was approximately 18.57 mV. As MA and DRP were negatively charged, a decrease in potential was observed after loading both of these onto the CeO2—NH2 surface, further indicating the successful synthesis of CeO2-DRP/MA. Considering the inherent Ce in CeO2 nanomaterials and the abundance of N and P in MA and DRP, these characteristic elements were detected to confirm the successful connection of MA and DRP with the nanomaterial surface. Uniform dispersion of CeO2-DRP/MA nanoclusters was achieved through ultrasonic technology, followed by imaging using HR-TEM and precise identification of elemental composition by analyzing the energy spectrum of specific elements. The experimental results (Fig. S1 in Supporting information) revealed the presence of three characteristic elements, Ce, N, and P, thereby confirming the successful synthesis of CeO2-DRP/MA nanomaterials and the effective connection between MA and DRP. The crystal structure was characterized by using powder X-ray diffraction (XRD) technology. XRD analysis results (Fig. 1e) showed that, compared with unmodified CeO2 nanoparticles, the diffraction patterns of the CeO2-DRP/MA nanoclusters modified with MA and DRP were not significantly different. This suggests that the introduction of MA and DRP did not affect the crystal structure of CeO2 nanomaterials after surface modification. This indicated that the CeO2-DRP/MA nanomaterials maintained their inherent crystal structure characteristics during biological functionalization. The modification density of MA and DRP on the surface of CeO2—NH2 was illustrated in Fig. S2 (Supporting information), the loading of MA and DRP on CeO2—NH2 was about 1.33 pmol/µg. CeO2-DRP/MA must cross the blood-brain barrier (BBB) to detect and treat AD. To examine the ability to cross the BBB, CeO2-DRP/MA nanomaterials were injected into mice through the tail vein. The mice were then euthanized at 12 and 24 h to collect brain tissue samples, which were used for fluorescence observations using an IVIS fluorescence imaging system. Owing to the DRP binding in the CeO2-DRP/MA nanomaterials to mitochondrial miRNA-204, the previously quenched fluorescent groups regained luminescence. Therefore, by measuring the fluorescence intensity changes in mouse brain tissue, the efficiency of nanomaterials in crossing the BBB could be directly evaluated. The experimental data (Fig. 1f) showed that over time, the fluorescence intensity in the mouse brain tissue gradually increased, indicating that the CeO2-DRP/MA nanomaterials could effectively penetrate the BBB and accumulate in brain tissue. Moreover, the distribution of CeO2-DRP/MA in peripheral organs further verified its ability of crossing BBB (Fig. S3 in Supporting information). In the design of CeO2-DRP/MA nanomaterials, CeO2 was chosen as the core component, because its nanoscale size was conducive to crossing the BBB as well as its significant reactive oxygen species scavenging ability. Reactive oxygen species scavenging potential was determined using 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) as a probe to detect reactive oxygen species. The experimental results (Fig. 1g) showed that compared with the control group, the green fluorescence and its intensity displayed by the CeO2-DRP/MA-treated cells were significantly reduced, suggesting that the CeO2-DRP/MA nanomaterial effectively cleared excess reactive oxygen species in AD model cells. To test the ability of CeO2-DRP/MA nanomaterials for detecting AD mitochondrial miRNA-204, CeO2-DRP/MA was added to a solution containing mitochondrial miRNA-204, and the fluorescence curve was observed within 1 h. The experimental results (Fig. 1h) showed that the fluorescence intensity of the FAM fluorescent groups in the nanomaterials gradually increased with increasing reaction time. The effectiveness of CeO2-DRP/MA nanomaterials in detecting mitochondrial miRNA-204 was confirmed, the fluorescence intensity of the FAM fluorescent groups increased with nanomaterial binding to the target mitochondrial miRNA-204, thereby achieving effective detection of AD-specific mitochondrial miRNA-204. To ensure that the CeO2-DRP/MA nanomaterials specifically reacted with mitochondrial miRNA-204, rather than with other mitochondrial miRNAs, their specificity was subsequently detected and analyzed. As shown in Fig. 1i, the CeO2-DRP/MA nanomaterials showed significant fluorescence recovery only in the presence of mitochondrial miRNA-204. In contrast, for the non-target miRNA-155 and miRNA-181, the FAM fluorescence intensity of the nanomaterial was comparable to that of the blank control group, confirming its highly specific detection ability for mitochondrial miRNA-204. To eliminate the potential impact of background signals on the fluorescence response, data standardization was performed using relative fluorescence intensity changes (F/F0), where F and F0 represented FAM fluorescence intensity in the presence or absence of target miRNA-204, respectively. As shown in Fig. 1j, a good linear relationship was found between the relative fluorescence intensity change (F/F0) and the logarithm of miRNA-204 concentration, covering the concentration range from 0.1 pmol/L to 1 nmol/L. The limit of detection (LOD) obtained by calculating three times the standard deviation of the blank control was 64.9 fmol/L, indicating the detection ability of CeO2-DRP/MA nanomaterials for mitochondria miRNA-204 at extremely low concentrations, demonstrating its potential application in trace biomarker detection.
After 24 h treatment with different concentrations of CeO2-DRP/MA, a cell counting kit-8 (CCK-8) assay was used to evaluate cell viability. As shown in Fig. 2a, the activity of SY5Y+Aβ cells gradually increased with the increase of CeO2-DRP/MA concentration, and the cell survival rate was restored to 98% when the CeO2-DRP/MA concentration reached 1 mmol/L. Further, the cytotoxicity of CeO2-DRP/MA in SY5Y normal cells and N2aSW AD model cells was extremely low and almost negligible. This indicated that CeO2-DRP/MA nanomaterials had good biocompatibility with normal cells and potential therapeutic effects on damaged nerve cells. After incubating SY5Y cells with CeO2-DRP/MA at a concentration of 1 mmol/L for 4 h, the detection ability of the nanomaterials for AD mitochondrial miRNA-204 was determined using confocal laser scanning microscopy (CLSM). As shown in Fig. 2b, strong green fluorescence from FAM was observed in SY5Y cells treated with Aβ, indicating a higher expression level of mitochondrial miRNA-204 in AD cells. The weak green fluorescence from FAM in normal cells indicated a significantly lower expression of mitochondrial miRNA-204 compared to that in AD cells. This phenomenon was also observed in N2aSW cells (Fig. S4 in Supporting information). These results confirmed that CeO2-DRP/MA exhibited good detection performance in cells. To test their mitochondria-targeting ability, co-localization of nanomaterials and mitochondria in cells was observed using CLSM. Mitochondria showed specific red fluorescence upon staining. In the presence of the target mitochondrial miRNA-204, the FAM fluorescent group in the CeO2-DRP/MA nanomaterials emitted green fluorescence. Therefore, by observing the colocalization of red and green fluorescence, it was possible to determine whether the CeO2-DRP/MA nanomaterials could successfully target and enter the mitochondria. As shown in Fig. 2c, the yellow region formed by the high overlap of red and green fluorescence indicates effective co-localization of CeO2-DRP/MA nanomaterials with mitochondria. Significant yellow co-localization signals were also observed in N2aSW cells (Fig. S5 in Supporting information), further confirming the targeting selectivity of CeO2-DRP/MA nanomaterials towards mitochondria in neural cells. Moreover, we detected the mitochondrial membrane potential of the SY5Y+Aβ cells treated with CeO2-DRP/MA. Compared to the negligible red-fluorescence-positive population in SY5Y+Aβ cells, strong red-fluorescence-positive cells appeared after being treated with the CeO2-DRP/MA (Fig. S6 in Supporting information). It validated the CeO2-DRP/MA could restore the damaged mitochondria via inducing remarkable change of potential of mitochondria.
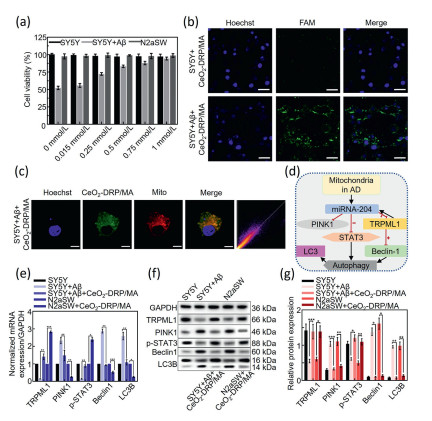
Figure 2
Figure 2.
(a) Viability of SY5Y and N2aSW cells treated with different concentrations of CeO2-DRP/MA (0–1 mmol/L) at 24 h. (b) CLSM images of different groups treated for 4 h (FAM: λex = 488 nm, λem = 520 nm; Hoechst: λex = 405 nm, λem = 420–480 nm), scale bars: 50 µm. (c) CLSM images of Aβ-treated SY5Y cells with CeO2-DRP/MA at 4 h (FAM: λex = 488 nm, λem = 520 nm; Mito: λex = 579 nm, λem = 599 nm; Hoechst: λex = 405 nm, λem = 420–480 nm). Scale bar: 20 µm. Pearson's value: 0.93. (d) Schematic illustration of gene regulation by CeO2-DRP/MA. (e) qRT-PCR analysis results showing TRPML1, PINK1, p-STAT3, Beclin1, and LC3B mRNA expression in different groups of treated cells. (f, g) Western blot analysis results for the expression of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B proteins in different groups of treated cells. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. Data in a, e, g are presented as mean ± SD (n = 3), and e, g are compared by one-way ANOVA. *P < 0.05, **P < 0.01.
TRPML1 is a negative feedback target gene for mitochondrial miRNA-204 and a target of the phosphatase and tensin homolog deleted on chromosome ten induced putative kinase 1 (PINK1) protein. It plays an important role in regulating the STAT3 signaling pathway. The known target proteins of PINK1 also include Aβ protein. TRPML1 participates in cellular autophagy regulation by directly affecting the STAT3 signaling pathway. Upon increased TRPML1 expression, STAT3 and Beclin-1 signaling molecules are activated in the cellular autophagy. This leads to STAT3 and Beclin-1 jointly responding and acting in regulating cellular autophagy (Fig. 2d). Therefore, by detecting the gene expression levels of STAT3, TRPML1, and Beclin-1, it is possible to explore how CeO2-DRP/MA nanomaterials treat AD. First, the effect of CeO2-DRP/MA nanomaterials on the cellular autophagy-mediated mechanism was observed at the mRNA level. The experimental results (Fig. 2e and Fig. S7 in Supporting information) showed that the mRNA expression levels of miRNA-204, PINK1, Beclin-1, and light chain 3 beta (LC3B) genes in the SY5Y+Aβ group cells treated with CeO2-DRP/MA were significantly reduced compared with those in the control group SY5Y+Aβ cells, whereas the mRNA expression levels of TRPML1 and STAT3 genes were significantly increased (P < 0.01), indicating that the entry of nanomaterials into cells inhibited the expression of mitochondrial miRNA-204, leading to increased expression of the negative feedback target gene TRPML1, which in turn promoted STAT3 expression. Upon receiving signals of high STAT3 and TRPML1 expression, the autophagy was regulated leading to a decrease in Beclin-1 expression, decreased expression of the cell autophagy marker gene LC3B. Similar changes in protein were also observed in N2aSW cells. The experimental results revealed that CeO2-DRP/MA entering cells effectively regulated cellular autophagy. After analyzing the regulation of cell autophagy by CeO2-DRP/MA at the mRNA level, its autophagy-regulated ability at the protein level was examined. As shown in Figs. 2f and g, the expression of PINK1, Beclin1, and LC3B proteins was significantly reduced in the CeO2-DRP/MA treatment group, whereas the expression of TRPML1 and p-STAT3 proteins was significantly increased compared with that in the control group (P < 0.01). This indicated that CeO2-DRP/MA inhibited the expression of mitochondrial miRNA-204 by targeting the mitochondria, leading to an increase in TRPML1 protein expression, which signaled an increase in STAT3 protein expression for downregulating Beclin-1 protein expression in the cellular autophagy. The decreased Beclin-1 protein expression further regulated cellular autophagy, leading to inhibition of LC3II conversion to LC3I in the autophagy-related protein LC3B. These experimental results further confirmed the ability of CeO2-DRP/MA to regulate cellular autophagy at the protein level.
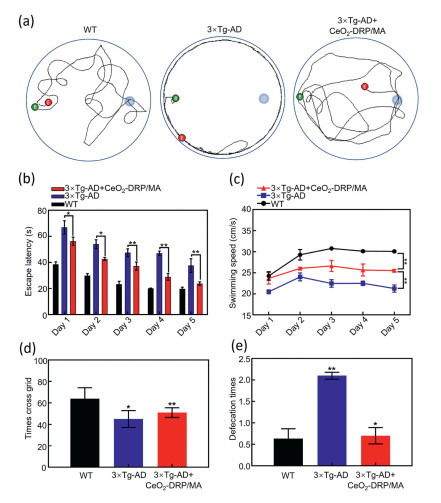
To observe whether the nanomaterials could restore cognitive function in mice, CeO2-DRP/MA was injected into their tail veins. All animal experiments were conducted in compliance with the regulations of the Animal Ethics and Welfare Committee (AEWC) of Longgang Central Hospital (No. 2024–008). The treatment period was set as two months, and medication was administered every two days. In water maze experiments, the swimming paths of mice were recorded in detail using high-definition cameras (Fig. 3a). The 3×Tg-AD model mice mainly mechanically swam along the edge of the water maze during the experiment but were unable to effectively locate the specific position of the previous platform. In contrast, the mice treated with CeO2-DRP/MA exhibited significantly purposeful swimming behavior, such as the ability to perform circular swimming within the target quadrant after the platform was removed. The mice in the CeO2-DRP/MA-treated group showed similarities in their ability to remember the past platform positions compared to that of the wild-type (WT) mice, indicating the effectiveness of CeO2-DRP/MA nanomaterials in alleviating cognitive dysfunction in 3×Tg-AD model mice. After observing the movement trajectory of the target platform-seeking behavior of the experimental group of mice, further experimental data were collected and analyzed to evaluate their memory and spatial learning abilities. During the training phase, the experimental group of mice searched for the quadrant of the previous target platform through free swimming, as shown in Fig. 3b. As the number of training days increased, the average time required for the mice in each experimental group to locate the platform gradually decreased. The decrease in the time required for the mice in the CeO2-DRP/MA-treated group to find the platform was statistically significant (P < 0.05). By calculating the average swimming speed of each group of mice reaching the platform during the training period, the results showed that the CeO2-DRP/MA-treated mice could locate the platform position faster than the control mice (Fig. 3c). The swimming times of each group of mice in the target platform quadrant during the testing period are shown in Fig. S8 (Supporting information). Compared with the 3×Tg-AD model mice, the WT and CeO2-DRP/MA-treated mice had a significantly longer residence time in the target quadrant. These results demonstrated the effectiveness of the CeO2-DRP/MA nanomaterials in restoring memory and spatial learning abilities in AD model mice, significantly improving their swimming speed, and enhancing their spatial localization ability. Furthermore, open-field test (OFT) experiments were used to explore the exploratory ability and anxiety status of the mice in each experimental group. The experimental results, as shown in Figs. 3d and e, showed that during the 5 min observation period, the number of times 3×Tg-AD mice crossed the cross grid was significantly reduced compared to that with WT mice and CeO2-DRP/MA-treated mice (P < 0.01), indicating that 3×Tg-AD mice exhibited relatively low exploration ability. Compared to WT mice, 3×Tg-AD mice exhibited more pronounced anxiety behaviors, such as frequent bowel movements. CeO2-DRP/MA not only significantly reduced the number of bowel movements in AD model mice as well as reduced their anxiety levels. Overall, CeO2-DRP/MA was found to effectively enhance the exploratory behavioral ability of 3×Tg-AD mice and alleviate their anxiety symptoms, further confirming the potential therapeutic effect of CeO2-DRP/MA nanomaterials in improving AD-related behavioral abnormalities.
Figure 3
Figure 3.
(a) Representative path tracings of WT mice, 3×Tg-AD mice, and CeO2-DRP/MA-treated 3×Tg-AD mice. (b) Escape latency of mice from different experimental groups. (c) Swimming speed of mice from different experimental groups. (d) Depression-related behaviors of mice from different experimental groups. (e) Anxiety-related behaviors of mice from different experimental groups. Data in b–e are presented as mean ± SD (n = 10) and are compared by one-way ANOVA. *P < 0.05, **P < 0.01.
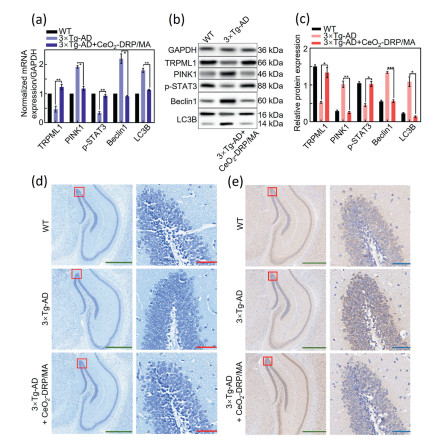
After animal behavioral experiments confirmed that CeO2-DRP/MA significantly improved cognitive impairment in AD mice, further exploration was conducted in 3×Tg-AD mice to determine whether CeO2-DRP/MA successfully mediated the expression of mitochondrial miRNA-204 and played an autophagy-regulated role in AD. The hippocampal tissue of each group of experimental mice was dissected, and total RNA was extracted. Quantitative real-time polymerase chain reaction (qRT-PCR) analysis confirmed that the mRNA expression of PINK1, Beclin1, and LC3B was significantly downregulated, whereas that of TRPML1 and p-STAT3 was upregulated following CeO2-DRP/MA treatment (Fig. 4a). Western blot analysis of mouse hippocampal proteins showed results consistent with those of qRT-PCR (Figs. 4b and c). In the brain tissues of mice treated with CeO2-DRP/MA, the expression levels of PINK1, Beclin1, and LC3B proteins were significantly lower than those in the control group, whereas those of TRPML1 and p-STAT3 proteins showed significant upregulation. These findings regarding both the mRNA and protein expression levels in mice demonstrated the effective miRNA-regulatory function of CeO2-DRP/MA. The Nissl staining experiment directly revealed the absence of neuronal cells (Fig. 4d). Compared to WT mice, significant neuronal nuclear contraction and low cell viability were observed in the hippocampus of 3×Tg-AD mice. However, hippocampal neurons in the CeO2-DRP/MA treatment group mice showed intact morphology, and their cell density did not differ from that of WT mice. To further evaluate the potential impact of CeO2-DRP/MA on the deposition of Aβ protein in the brain of 3×Tg-AD mice, the distribution and content of Aβ protein in the hippocampus region of the mouse brain were examined using immunohistochemical and Aβ42/40 enzyme-linked immunosorbent assay (ELISA) kits. The experimental results are shown in Fig. 4e and Fig. S9 (Supporting information). The hippocampus of 3×Tg-AD mice exhibited muti-directional Aβ protein expression. After treatment, with CeO2-DRP/MA, the number and size of protein deposits were reduced. At the meantime, Fig. S10 (Supporting information) demonstrated the positive safety of CeO2-DRP/MA in mice.
Figure 4
Figure 4.
(a) qRT-PCR analysis of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B mRNA expression in the hippocampus of mice from different experimental groups. (b, c) Western blot analysis of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B protein levels in the hippocampus of mice from different experimental groups. (d) Nissl staining of mouse hippocampus (green scale bars: 500 µm; red scale bars: 50 µm). (e) Immunohistochemical analysis of Aβ expression in the mouse hippocampus (green scale bars: 500 µm, blue scale bars: 50 µm). Data in a, c are presented as mean ± SD (n = 3) and are compared by one-way ANOVA. *P < 0.05, **P < 0.01.
These results demonstrated that CeO2-DRP/MA effectively reduced the degree of neuronal cell damage in AD mice, confirming that CeO2-DRP/MA exerts positive therapeutic effects on AD in vivo.
In summary, we designed a mitochondria-targeted detection system CeO2-DRP/MA to diagnose AD and treat damaged mitochondria. CeO2-DRP/MA nanomaterials can cross the BBB and utilize the unique reactive oxygen species clearance mechanism of CeO2 for AD treatment. Owing to the ability of mitochondrial aptamers to target the mitochondria, CeO2-DRP/MA nanomaterials have successfully targeted the mitochondria both in vitro and in vivo. The DNA-RNA hybrid chains in CeO2-DRP/MA allowed the detection of mitochondrial miRNA-204. Simultaneously, by silencing the expression of mitochondrial miRNA-204, the mitochondrial membrane potential was restored to normal levels. The performance of CeO2-DRP/MA in the simultaneous detection of mitochondrial miRNA-204 and treatment of AD thus provides a prospective research approach for the development of nanodiagnostic and therapeutic platforms for targeting mitochondrial miRNAs.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This work was supported by National Key Research and Development Program of China (No. 2023YFD1800105), Long-gang District Medical and Health Technology Research Project (No. LGKCYLWS2022024), the Non-profit Central Research Institute Fund of Chinese Academy of Medical Sciences (No. 2023-JKCS-12), Science and Technology Research Project of Colleges and Universities in Hebei Province (No. ZD2020306) and the Medical Health Technology Innovation Project of Chinese Academy of Medical Sciences (No. 2021-I2M-1-058).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2024.110614.
[1]
C. Reitz, M. Vance, T. Foroud, et al., Nat. Rev. Neurol. 19 (2023) 261–277. doi: 10.1038/s41582-023-00789-z
A. Lee, C. Kondapalli, D.M. Virga, et al., Nat. Commun. 13 (2022) 4444.
[15]
C. Insup, W. Ming, Y. Seungyeul, et al., Nat. Cell Biol. 25 (2023) 963–974.
[16]
L. Yi, Y. Peng, M. Ran, et al., Acta Pharm. Sin. B. 14 (2024) 1380–1399. doi: 10.3390/mi15111380
[17]
Z. Wei, X. Cheng, S. Ji, et al., Acta Pharm. Sin. B 12 (2022) 1019–1040. doi: 10.3390/sym14051019
[18]
Q. Yang, C. Yan, Y. Sun, et al., Adv. Sci. 23 (2024) 2400480.
[19]
A.N. Ralph, W. Jerzy, K. Asok, et al., J. Neuropath. Exp. Neurz. 64 (2005) 113–122.
[20]
M. Luo, Y. Pang, J. Li, et al., Acta Pharm. Sin. B 14 (2024) 635–652.
[21]
G.E. Stutzmann, R. Marr, S.H. Mustaly, S. Schrank, Alzheimers Dement. 17 (2021) e057599.
[22]
D. Tosun, R. Ellis, M.W. Weiner, et al., Alzheimers Dement. 17 (2021) e053246.
[23]
S. Minkyoung, S. Soo, K. Kyung, et al., Biomedicines 9 (2021) 1764.
[24]
G. Prashanth, R. Hemachan, K. Subodh, Ageing Res. Rev. 73 (2022) 101529.
[25]
S. Dolores, J. Alfredo, R. Jose, et al., Cells 10 (2021) 113.
[26]
J. Long, Z. Min, Y. Jian, et al., BMC Med. 19 (2021) 264.
[27]
R. Arun, C. Chandra, H. Ken, ACS Sens. 6 (2021) 3176–3181.
[28]
L. Agosta, D. Arrieta, M. Dzugutov, et al., Angew. Chem. Int. Ed. 62 (2023) e202303910.
[29]
J. Xu, Y. Wang, K. Wang, et al., Angew. Chem. Int. Ed. 62 (2023) e202302877.
[30]
C. Xiang, Z. Ling, W. Ke, et al., Chin. Chem. Lett. 35 (2024) 108461.
[31]
A. Lorenzo, D. Arismendi, D. Mikhail, et al., Angew. Chem. Int. Ed. 62 (2023) e202303910.
[32]
K. Jenkinson, M.C. Spadaro, V. Golovanova, et al., Adv. Mater. 35 (2023) 2306447.
[33]
A. Yoko, Y. Omura, K. Ninomiya, et al., J. Am. Chem. Soc. 146 (2024) 16324–16331. doi: 10.1021/jacs.4c05106
[34]
X. Zhou, Q. Zhou, Z. He, et al., Nanomicro Lett. 16 (2024) 156.
[35]
X. Xu, Z. Han, D. Li, et al., Adv. Funct. 34 (2024) 2313198.
Figure 1
(a) Synthetic route of CeO2-DRP/MA and schematic diagram of CeO2-DRP/MA use in miRNA-204 biomarker detection and AD therapy. (b) HR-TEM image of CeO2-DRP/MA. Scale bar: 20 nm. (c) Size analysis of CeO2—NH2 and CeO2-DRP/MA. (d) Zeta potential analysis of CeO2—NH2 and CeO2-DRP/MA. (e) X-ray diffraction patterns of CeO2—NH2 and CeO2-DRP/MA. (f) Fluorescence imaging of 3×Tg-AD mouse brains at various time points after CeO2-DRP/MA injection. (g) Active oxygen removal ability based on CLSM images of different group cells incubated with CeO2-DRP/MA for 24 h (λex = 495 nm, λem = 529 nm). Scale bar: 100 µm. (h) Fluorescence intensities of CeO2-DRP/MA in the presence of miRNA-204 in solution at different time intervals. (i) The detection specificity of CeO2-DRP/MA for mitochondrial miRNA-204. (j) The detection sensitivity of CeO2-DRP/MA for mitochondrial miRNA-204. Data in c, i are presented as mean ± standard deviation (SD) (n = 3).
Figure 2
(a) Viability of SY5Y and N2aSW cells treated with different concentrations of CeO2-DRP/MA (0–1 mmol/L) at 24 h. (b) CLSM images of different groups treated for 4 h (FAM: λex = 488 nm, λem = 520 nm; Hoechst: λex = 405 nm, λem = 420–480 nm), scale bars: 50 µm. (c) CLSM images of Aβ-treated SY5Y cells with CeO2-DRP/MA at 4 h (FAM: λex = 488 nm, λem = 520 nm; Mito: λex = 579 nm, λem = 599 nm; Hoechst: λex = 405 nm, λem = 420–480 nm). Scale bar: 20 µm. Pearson's value: 0.93. (d) Schematic illustration of gene regulation by CeO2-DRP/MA. (e) qRT-PCR analysis results showing TRPML1, PINK1, p-STAT3, Beclin1, and LC3B mRNA expression in different groups of treated cells. (f, g) Western blot analysis results for the expression of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B proteins in different groups of treated cells. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. Data in a, e, g are presented as mean ± SD (n = 3), and e, g are compared by one-way ANOVA. *P < 0.05, **P < 0.01.
Figure 3
(a) Representative path tracings of WT mice, 3×Tg-AD mice, and CeO2-DRP/MA-treated 3×Tg-AD mice. (b) Escape latency of mice from different experimental groups. (c) Swimming speed of mice from different experimental groups. (d) Depression-related behaviors of mice from different experimental groups. (e) Anxiety-related behaviors of mice from different experimental groups. Data in b–e are presented as mean ± SD (n = 10) and are compared by one-way ANOVA. *P < 0.05, **P < 0.01.
Figure 4
(a) qRT-PCR analysis of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B mRNA expression in the hippocampus of mice from different experimental groups. (b, c) Western blot analysis of TRPML1, PINK1, p-STAT3, Beclin1, and LC3B protein levels in the hippocampus of mice from different experimental groups. (d) Nissl staining of mouse hippocampus (green scale bars: 500 µm; red scale bars: 50 µm). (e) Immunohistochemical analysis of Aβ expression in the mouse hippocampus (green scale bars: 500 µm, blue scale bars: 50 µm). Data in a, c are presented as mean ± SD (n = 3) and are compared by one-way ANOVA. *P < 0.05, **P < 0.01.

 DownLoad:
DownLoad:

 下载:
下载:





