引用本文:
王子健, 杨静波, 李光谱, 孙宁宁, 孙万春, 彭其胜, 刘宁. 低浓度甲醛对多肽和蛋白化学修饰的质谱研究[J]. 分析化学,
2016, 44(8): 1193-1199.
doi:
10.11895/j.issn.0253-3820.160139 Citation:
WANG Zi-Jian, YANG Jing-Bo, Li Guang-Pu, SUN Ning-Ning, SUN Wan-Chun, PENG Qi-Sheng, LIU Ning. Chemical Modifications of Peptides and Proteins with Low Concentration Formaldehyde Studied by Mass Spectrometry[J]. Chinese Journal of Analytical Chemistry,
2016, 44(8): 1193-1199.
doi:
10.11895/j.issn.0253-3820.160139
Key Laboratory of Zoonosis, Ministry of Education, Jilin University, Changchun 130062, China
Received Date:
29 February 2016 Revised Date:
17 June 2016
Fund Project:
This work was supported by the National Natural Science Foundation of China (Nos. 81472030, 31372409, 21175055)
Abstract:
Formaldehyde has been widely employed to immobilize clinical tissue specimens, inactivate toxins and viruses in biomedical fields. Formaldehyde can react with active groups in bio-molecules such as proteins, resulting in protein cross-linking, inactivation, and immobilization. By using several standard peptides and tryptic peptides from matrix protein of influenza virus as experimental models, we studied the chemical modifications of peptides and proteins with formaldehyde by matrix-assisted laser desorption ionization time-of-flight mass spectrometry and nano-electrospray quadruple time-of-flight tandem mass spectrometry. The reaction between formaldehyde and peptides was performed under the same conditions as those during inactivation of virus (4℃, 0.025% Formalin (V/V), 37% formaldehyde solution (w/w), and 72 h). The results indicated that under above conditions, formaldehyde could react with amino group of N-terminus of standard peptide to generate a methylol adduct, which was further condensed into an imine to generate +12 Da product. Besides, formaldehyde could react with side chain of two amino acids such as arginine and lysine, yielding +12 Da product respectively. The analysis of the reaction between formaldehyde and tryptic peptides from matrix protein of influenza virus showed that +24 Da products could be detected in most peptides due to combinational contribution from N-terminus of peptide (+12 Da) and side chain of C-terminal arginine or lysine (+12 Da). Moreover, a +36 Da product was detected for a peptide with miss-cut site. The results indicated that low-concentration formaldehyde primarily reacted with amino group on N-termini of peptides and proteins, as well as the side chains of arginine and lysine residues. The present study suggested an effective mass spectrometry-based method for analyzing the reaction between low-concentration formaldehyde and peptides and proteins, thus provided strategies for interpretation for the mass spectra of reaction products.
分别将多肽样品溶解于50 mmol/L磷酸盐缓冲液(pH 7.5),使其终浓度为66μmol/L。将福尔马林用水稀释200倍,取2.5μL加入50μL多肽样品溶液中,迅速振荡混合均匀,在4℃放置一定时间后,用C18 Zip Tip吸头脱盐,点靶,与基质溶液混合并在室温干燥后,进行MALDI-TOF质谱分析。另取脱盐的样品适量,进行ESI质谱分析。
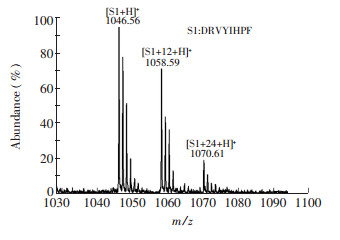
Figure 1.
Matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS) analysis of reaction between peptide S1 and formaldehyde
Figure 2.
Nano-electrospray quadruple time-of-flight tandem mass spectrometry (NESQ-TOF-MS/MS) analysis of peptide S1 with m/z 523.76 (A) and two products with either m/z 529.76 (B) or m/z 535.76 (C) formed during incubation of S1 with formaldehyde
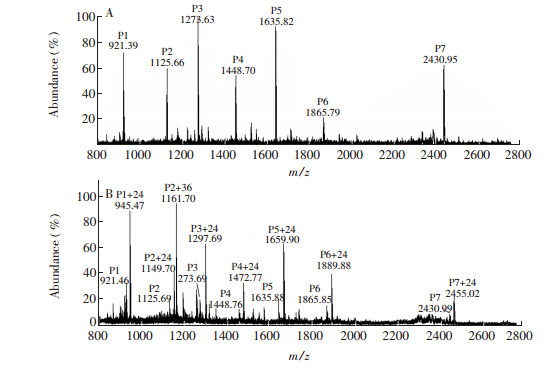
Figure 3.
(A) MALDI-TOF-MS analysis of tryptic peptides from matrx protein of influenza virus. (B) MALDI-TOF-MS analysis of tryptic peptides from matrx protein of influenza virus, which were incubated with low concentration of formaldehyde
Fraenkel-Conrat H,Olcott H S.J.Am.Chem.Soc.,1948,70(8):2673-2684
[13]
Fraenkel-Conrat H,Olcott H S.J.Biol.Chem.,1948,174(3):827-843
[14]
Consden R,Gordon A H,Martin A J.Biochem.J.,1946,40(4):580-582
[15]
Metz B,Kersten G F,Hoogerhout P,Brugghe H F,Timmermans H A,de Jong A,Meiring H,ten Hove J,Hennink W E,Crommelin D J,Jiskoot W.J.Biol.Chem.,2004,279(8):6235-6243
[16]
Pappas C,Matsuoka Y,Swayne D E,Donis R O.Clin.Vaccine Immunol.,2007,14(11):1425-1432
[17]
Liu N,Song W,Lee K C,Wang P,Chen H,Cai Z.J.Am.Soc.Mass Spectrom.,2009,20(2):312-320
Figure 1
Matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS) analysis of reaction between peptide S1 and formaldehyde
Figure 2
Nano-electrospray quadruple time-of-flight tandem mass spectrometry (NESQ-TOF-MS/MS) analysis of peptide S1 with m/z 523.76 (A) and two products with either m/z 529.76 (B) or m/z 535.76 (C) formed during incubation of S1 with formaldehyde
Figure 3
(A) MALDI-TOF-MS analysis of tryptic peptides from matrx protein of influenza virus. (B) MALDI-TOF-MS analysis of tryptic peptides from matrx protein of influenza virus, which were incubated with low concentration of formaldehyde


 下载:
下载:


 下载:
下载:




