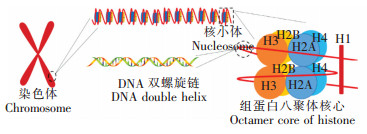
图 1
染色体、核小体、组蛋白及DNA示意图
Figure 1.
Chromosome, nucleosome, histone and DNA

基因组的完整性在生命活动过程中一直会受到各种威胁,包括DNA单链断裂(SSBs)、DNA双链断裂(DSBs)、DNA和DNA交联、DNA与蛋白质交联、DNA碱基加合、DNA置换、DNA移码突变等。引起这些损伤的原因既包括内源因素,如细胞代谢,也有外在因素,如环境中有害化学物质或者各种辐射的暴露[1]。在这些损伤中,DNA双链断裂最为严重,因为其可以导致基因组重排、遗传信息丢失或者细胞死亡[2]。这种分子和细胞上的微观变化,最终有可能引起有机体宏观上的伤害,如肿瘤发生等。因此,DSBs的检测对评估致DNA双键断裂因素以及由此可能引起的不良后果具有重要意义。DSBs的检测技术通常依赖于特异的DNA双链断裂后的信号响应分子或者修复通路信号分子。在大部分的真核生物中,DSBs可通过Ku异质二聚体及Mrell-Rad50-Xrs2(MRX)蛋白复合体被检测[2]。然而,自Rogakou于1998年首次发现并阐明γH2AX可以作为细胞中一种快速而灵敏的DSBs应答信号分子以来[3],人们已经对至少200多种基于γH2AX的化学物质的基因毒性进行了评估。目前,与γH2AX相关理论研究臻于完善,检测技术灵活多样,一些方便、快速和准确的检测技术已经在毒理学检测和评估、临床医药学等各个方面得到了广泛应用。
真核生物染色质的基本结构单位为核小体,核小体由DNA和组蛋白构成,每个核小体单位包括200 bp左右的DNA和一个组蛋白八聚体以及一分子的组蛋白H1,组蛋白八聚体由各两分子的H2A、H2B、H3和H4组成(如图 1)。人类基因组中除H2A外,还有其它5种H2A基因,包括20世纪80年代鉴定的H2AX、H2AZ[4],90年代发现的macroH2A1和macroH2A2[5, 6],以及本世纪初发现的H2A-Bbd[7]。此外,20世纪80年代,人们还在酵母中发现了H2A1和H2A2两种变体[8]。所有这些H2A变体的数量虽然非常少,但是在染色质代谢过程中可能扮演着非常重要的角色。如H2AX在DSBs修复过程中具有非常突出和重要的作用。
H2A变体和H2A的序列在很多位置都具有较大差异,然而H2AX与其主要的差异表现在多肽链的羧基末端,而且从酵母到人类,其序列高度保守[9]。在不同生物中,H2AX约占H2A含量的2%~10%[3],其编码基因位于11q23.2-q23.3,共编码142个氨基酸,其中139-140位氨基酸形成了一个保守的丝氨酸-谷氨酰胺(Ser-Gln,SQ)的结构域(如图 2)。该结构域中的Ser残基可以被磷脂酰肌醇-3-激酶(phospatidylinositol 3-kinase,PI3K)家族的运动失调性毛细血管扩张症突复基因(Ataxia Telangiectasia Mutated gene,ATM)、ATM和Rad3相关蛋白(ATM and Rad3-related protein,ATR)以及DNA依赖性蛋白激酶催化亚单位(DNA-dependent Protein Kinase catalytic subunit,DNA-PKcs)等磷酸化[10],其磷酸化后的产物被称为γH2AX。

很多因素可以导致细胞内γH2AX的形成,包括紫外线[11]、X-射线[12]、热效应[13]、带电重离子[14]、低氧[15]、高渗透压[16]、低pH[17]等,以及DNA损伤试剂(如Bleomycin[18]及Etoposide[19]等)、DNA插入试剂(如Cisplatin[20])、DNA/RNA合成抑制剂(如Hydroxyurea[21]及Actinomycin D[22])、重金属(如镉[19])、产活性氧试剂(如过氧化氢[23])以及复杂化学混合物等化学试剂,还包括诸如细胞凋亡[24]、有丝分裂[25]、衰老[26]及微生物[27]作用等生物因素。细胞为了应答这些因素产生的DSBs,修复染色质结构,维护基因组的稳定性,于DSBs处招募大量信号分子和修复蛋白,对DNA进行修复,而γH2AX焦点的形成就是DSBs发生后早期被招募的分子复合体之一。研究表明,细胞在受到电离辐射后3 min内就可检测到γH2AX,10~30 min就可达到高峰[28]。
H2AX的磷酸化主要由ATM和DNA-PKcs催化产生,但是两者的作用具有一定差异。其中,ATM被认为是H2AX磷酸化的主要实施者(主要过程见图 3),当细胞受到电离辐射而发生DSBs后,ATM与Nbs1相互作用,并从细胞质转移至细胞核内,在细胞核内的DSBs位点使H2AX磷酸化,而且NBs1羧基末端与ATM相互作用的结构域在ATM有效的磷酸化过程中是必须的。当NBs1羧基末端与ATM缺少其一,H2AX磷酸化主要由DNA-PKcs催化完成[29]。DNA-PKcs是一种核内丝氨酸/苏氨酸激酶,由一个催化亚基和一个DNA结合的Ku70/80异质二聚体构成[30]。有研究报道,DNA-PKcs被抑制后,电离辐射诱导的γH2AX水平显著降低,同时在ATM缺失时,仍能检测到高水平的γH2AX,而一旦ATM和DNA-PKcs都缺失时,γH2AX水平显著降低,说明DNA-PKcs在H2AX的磷酸化中是必须的且具有一定的主导作用[31]。然而,文献[30]表明,DNA-PKcs被抑制后,ATM的活性增强,γH2AX数量增加,说明ATM是γH2AX形成的关键激酶。虽然上述两种结论并不一致,但是至少说明DNA-PKcs在H2AX磷酸化形成γH2AX时具有重要作用,但普遍的研究认为其仅是对ATM催化H2AX磷酸化形成γH2AX的一种补充作用[10]。此外,除了ATM和DNA-PKcs外,PI3K家族成员的ATR被认为是内源因素诱导形成γH2AX的主要磷酸化激酶,如细胞复制时有丝分裂过程中形成的γH2AX主要由ATR催化产生[32]。

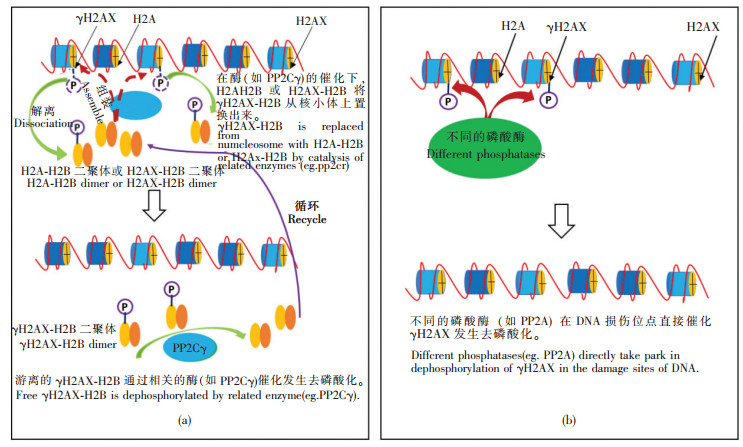
目前,虽然γH2AX形成的分子机制及其在DNA损伤修复中的作用已经获得了充分的认识,但是从染色体上移除以及恢复为H2AX的分子机制还有待于进一步探索。因为,目前仅除了发现一些去磷酸化的酶外,详细的分子信号通路尚处于初步研究阶段。据推测γH2AX的去磷酸化可能有两种途径实现,一是通过组蛋白交换方式从染色体上解离(如图 4a),另一种是以磷酸酶为主而介导的γH2AX去磷酸化[10](如图 4b),而且可能以后一种为主,因为目前已经发现了多种催化γH2AX去磷酸化的磷酸酶,如Pph3,PP2A,PP4,PP6,PP2Cγ,Wip1(wild-type p53-induced phosphatase 1)等。虽然不断发现新的磷酸酶可用于DSBs修复后γH2AX的去磷酸化,但是关于这些酶在γH2AX的去磷酸化过程中的详细分子机制,如催化反应的调控、产生时间控制,详尽的信号通路等尚处于初步的研究阶段。
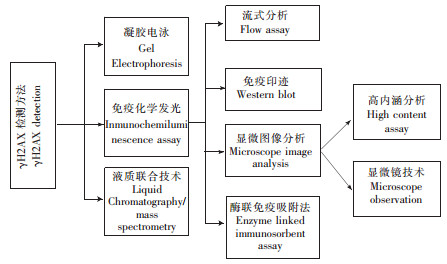
自1998年Rogakou首次分离并发现γH2AX后[3],γH2AX的检测方法不断得到发展和改进。先后有免疫印迹(Western blot,WB)、显微图像分析技术(Microscope image analysis,MIA)、酶联免疫吸附分析(Enzyme linked immunosorbent Assay,ELISA)、流式分析(Flow assay,FA)、高内涵(High content assay, HCA)及液相色谱-质谱联用(Liquid chromatography/mass spectrometry,LC/MS)等技术用于检测和分析γH2AX,各主要检测方法如图 5。
凝胶电泳(Gel electrophoresis,GE)是分离生物大分子的一种常用色谱分析方法,γH2AX最初的发现主要依赖于双向凝胶电泳和同位素标记技术的联用[3],能够直接将γH2AX从多种组蛋白中分离出来,可进行绝对定量分析。虽然该方法是发现γH2AX的主要技术,但是所需样本量较大,过程比较繁琐和费时,检测通量不高。
最近,日本科学家Shun Matsuda创建了一种可绝对定量检测γH2AX的LC/MS方法[48]。该技术分别以源于H2AX和γH2AX的ATQASQEY和ATQA(pS)QEY多肽片段以及对应的同位素标记多肽为检测目标,建立了LC/MS检测的相应条件。提取细胞中的组蛋白,采用胰蛋白酶进行梯度消解,使H2AX和γH2AX分别释放出ATQASQEY和ATQA(pS)QEY,通过已建立的LC/MS方法进行检测。经检测,10μmol/L喜树碱给药1 h的HeLa细胞中γH2AX的绝对含量为6.2×105分子/细胞,而H2AX和γH2AX的含量之和达到3.3×106~3.6×106分子/细胞[48]。虽然该方法具有可绝对定量、广泛的适应性和高通量测定等优点[48],但是前处理较为费时,其准确性、稳定性和灵敏性还需要进一步的验证。
流式分析依赖于流式细胞仪可以对单个细胞进行检测,因此可利用荧光抗体检测细胞内的γH2AX。Smart等[35]通过流式细胞仪检测细胞内的γH2AX评价有毒物质的基因毒性,发现其对Ames test的预测灵敏性达到100%,特异性达到67%,一致性达到82%,并对体外基因毒性哺乳动物实验的预测灵敏性达到91%,特异性性达到89%,一致性达到91%。流式细胞仪是目前测定γH2AX的一种非常有用且被广泛接受的技术。虽然该技术在实际中得到了广泛应用,但是其只能测定细胞内γH2AX的荧光强度,并不能进行测定细胞内的γH2AX焦点数量与分布,而且检测的灵敏性比显微镜法低[36]。此外,该技术对上样量具有一定的要求,样品检测通量并不及酶联免疫吸附分析法高,但比传统的免疫印迹技术操作简单、检测速率高。
荧光显微技术是最直观也是最方便的检测γH2AX的技术之一,可以直接通过荧光显微镜对细胞内的γH2AX焦点进行计数,并对其荧光强度的大小进行定性评判,从而比较不同样品处理后细胞内的γH2AX含量。如100μmol/L H2O2可以使CaCo-2细胞在处理后24 h产生18.01个γH2AX焦点,而300μmol/L啶虫脒在同样条件下只产生了8.83个γH2AX焦点[39]。该方法直观、简单、无需特殊的贵重仪器。但缺点也比较突出,主要包括人为计数荧光焦点和判断荧光强度的主观性较强,相对定量分析的准确度不高等。随着图像分析技术和人工智能的发展,目前可使用图像分析软件或者特殊仪器对结果进行分析和计算,减小了人为误差的同时大大节省了时间,提高了检测准确性。如计算机图像辅助软件ImageJ、NIH Image、MetaMorph、Image Pro等的使用极大地提高了计数通量,减少了主观性和潜在的计数偏好。虽然这些弥补了很多人工计数的缺点,但也引入了诸如单个细胞核识别、分析参数值的设定没有参考标准等问题[40]。为解决这些问题,不断有新的软件或系统平台被开发出来,如自动焦点分析软件、AKLIDES platform系统等的应用实现了γH2AX分析检测的自动化,而且分析检测通量明显提高[39-42]。
免疫印迹又称为蛋白质印迹或Western Blot, 是在凝胶电泳和固相免疫技术上发展起来的一种新的分子免疫印迹技术。免疫印迹技术准确,稳定可靠,使用较广,可对目标蛋白质进行定性和半定量分析,结合化学发光技术可对多个样品中的目标蛋白表达量进行比较,且得到的蛋白还可进行其它方面的进一步分析。通常,免疫印迹需要将目标蛋白提取和分离,操作较为繁琐,不利于大规模的筛选分析。随着检测仪器的发展,In-cell western technology (ICW)技术极大地提高传统的免疫印迹的检测通量和检测速率。Khoury等[34]对基于ICW的γH2AX检测评价基因毒性的高通量筛选实验进行详细研究,发现基于ICW的γH2AX检测对基因毒性化合物的敏感性为75%,特异性达到95%~100%,而且可以高通量检测γH2AX。ICW技术不仅极大地提高了传统技术的检测速率,同时也弥补了传统方法检测通量较低的缺点,但是ICW技术的使用需要借助特殊的光学显色扫描仪器,且不能进行绝对定量分析。此外,该技术不能获得对细胞内γH2AX焦点数目和分布的详细信息。
高内涵分析(HCA)是在荧光显微镜技术上不断发展中产生的,最初是Cellomics公司为解决高通量药物筛选而研发的[43]。HCA是一种在保持细胞结构和功能完整性的前提下,应用高分辨率的荧光数码系统从三维立体层面同时检测被检样品对细胞的诸如细胞形态、生长分化状态、迁移与凋亡、细胞代谢途径以及信号通路等生理状态和属性影响的高通量筛选手段[44]。Carolina等[45]对采用HCA检测γH2AX的体外基因毒性实验进行了评价,发现基于HCA的体外γH2AX实验的准确度达到86%,敏感性达到86%~92%,特异性达到80%~88%,说明HCA可以有效检测细胞内的γH2AX。HCA可以使用自动显微镜获取细胞图像,并能通过图像分析软件测定不同参数,如焦点亮度、每个细胞内的焦点数目及分布等;还可以对生成的图像进行储存,以便再次分析或者质量控制检查等[1]。此外,HCA在高通量和敏感性上对微孔板评分和检测γH2AX更具优势,因为它是在96孔或384孔板进行分析,与流式细胞术检测相比,HCA所需细胞数量和试剂更少,检测通量更大,且同一实验获得的信息量更多[46]。目前,HCA的发展仍存在一些不足:(1)与流式细胞仪不同,HCA仅能根据样本的准备情况,在一定的荧光强度、激发光与发射光波长范围等限制条件下使用,所以尚未得到产业化规模的应用;(2)多数HCA仪器不具有比例成像分析和分析细胞内所有荧光物质的能力,各种荧光染料的整合应用以及将成像转化成HCA模式以供分析仍有不足,成像技术有待进一步发展[47]。
在酶联免疫吸附分析法(ELISA)中,利用酶标抗体识别γH2AX,再催化特异性底物发光,最后通过发光强度对γH2AX进行半定量分析。由于可采用酶标仪和酶标板进行高通量测定,因此也被广泛用于γH2AX的测定。2010年,Kaori等[37]首次采用该方法对γH2AX进行了检测,无需破坏细胞,可直接对细胞内的γH2AX进行测定,测定通量高、操作方便简单,可采用96孔板一次对4种物质的10个浓度进行平行测定,且对测试物质所需量很低,比荧光显微图像分析法定量准确性好,比流式分析便捷。本实验室也曾对该方法的测量条件进行优化,并对卷烟烟气的基因毒性进行了评价[38]。
γH2AX可以作为DSBs的一种生物标志物, 且γH2AX焦点的形成和DSBs的数量存在1:1的关系[49]。因此,可以通过γH2AX检测评估环境毒物和致癌化学试剂的基因毒性。目前,已经通过γH2AX实验对200多种化学物质的基因毒性进行了评价,这些物质既包括卷烟烟气这样的复杂混合物,也包括多环芳烃等这样的单体化合物,以及各种辐射等。与传统的体外基因毒性方法相比,如鼠伤寒沙门氏菌实验(Ames test)、小鼠淋巴瘤实验、染色体畸变实验、微核实验、彗星实验、绿屏实验及酵母DEL实验等,γH2AX实验具有通量高、灵敏性高和假阳性报出频率低等优点,可以作为一种强有力的基因毒性评价的预筛选实验。如Yu等[50]通过彗星实验和γH2AX实验对MNNG(N-methyl-N’-nitro-N-nitroso-guanidine)导致的DNA损伤进行了比较研究,发现中性彗星实验和γH2AX虽然均能在低浓度下检测出DNA损伤,但中性彗星实验在高浓度的MNNG中未能检测出细胞内的DNA损伤。再如,以原核微生物为检测模型的Ames实验可能对一些仅能致真核生物产生基因毒性的化合物的检测能力是有限的,且具有抗生素类性质的化合物也是明显不适于Ames实验[1]。
尽管γH2AX实验展现出了诱人的体外基因毒性评价前景,但是在实际应用过程中仍有一些问题需要注意。(1)细胞种类的选择合适的细胞系对γH2AX实验的敏感性影响很大,有研究者认为,正常表型细胞比肿瘤细胞更为可靠,这是因为肿瘤细胞的DNA损伤修复机制和细胞周期调控功能均发生了异常变化[51, 52],而γH2AX参与细胞应答DNA损伤和细胞周期。例如,在无DSBs存在的肿瘤细胞内发现了γH2AX,进而推测这可能仅是一种对“致瘤压力”的应答[45]。同时,许多前毒性化合物需要代谢激活后才能表现出毒性,而大部分正常表型细胞和肿瘤细胞内都不存在相应的代谢激活酶系,因此需要加入外源的激活酶系,如鼠肝S9等,但是这些酶系的添加只能作为短时处理,因为它们在体外的活性有限,且对细胞具有一定的毒性。虽然这些困难可以通过使用其它代谢激活系统,如人肝微粒体或者含有代谢系统的细胞(如HepaRG和干细胞等)改善,但是这些并不能完全解决问题,例如使用HepaRG细胞系测试对呼吸系统能产生毒性的化学物质可能是不合适的[25, 53]。(2)细胞周期由于γH2AX参与细胞周期循环,因此细胞周期对γH2AX分析具有一定的影响,Markus等[54]指出,低比例的G2期细胞可以显著增大分析误差。(3)γH2AX可能并不完全代表DSBs越来越多的研究表明,γH2AX并不是一个非常明确的DSBs标志物,γH2AX之所以目前广泛被认为是DSBs的标志物,可能存在过分的解读或者误导[52-57]。所以,γH2AX实验用于毒理学上的研究还需要进一步研究。(4)γH2AX焦点的消失和DNA的损伤修复可能并不完全相关目前,人们并不清楚可视化的γH2AX焦点的消失和最终DSBs的修复是否相关,但是在实际研究过程中,只要认真考虑到这点,并不能否认γH2AX能够被利用去分析DSBs的修复过程[54]。
γH2AX及其检测不仅对毒理学研究和应用具有很大影响,而且对临床医药学上的研究和应用的影响也很大。主要表现在以下几个方面:(1)疾病的诊断复制压力引起的DNA双链断裂水平的提高不仅见于肿瘤中,而且也见于癌前病变[58]。因此,通过检测机体组织中的γH2AX水平就可以得知细胞中DNA双链断裂的情况,以辅助癌前病变的检查、肿瘤的早期发现和治疗[59]。Mah等[60]指出,γH2AX还可以作为衰老和体内慢性炎症的分子标志,因此,通过检测特定样品中的γH2AX可以评估机体衰老进程和可能存在的慢性炎症的程度。(2)药效或疗效评价大多数抗肿瘤药物(如博来霉素,喜树碱和依托泊苷等)或者治疗手段(如放疗及化疗等)基于基因毒性应激作用,可严重损伤肿瘤细胞DNA或干扰其代谢,最终使肿瘤细胞坏死或者凋亡。而γH2AX可以很好地监测细胞内的DNA损伤情况,因此通过分析γH2AX的水平可以指导医师及时地对患者的用药反应进行评价,如用药种类或给药剂量等[61]。如Kinders等[62]提出γH2AX可以作为拓扑异构酶I抑制剂活性的药效标志。(3)药物研发和临床试验通过分析和检测,γH2AX可以研究药物作用机理或者评估新药药效。如利用γH2AX分析可以研究调节细胞生长抑制剂的多重耐药性问题[63, 64],还可以对临床新药的毒性进行评估,如帕比司他对前列腺癌的治疗,冬凌草甲素对胰腺癌的治疗等[58]。
自γH2AX被发现至今不过十余年,在理论研究和实际应用上取得了非常多的成就。理论上,其形成主要是受PI3K家族的蛋白激酶催化,其去磷酸化主要依赖于细胞内的诸如PP2A,PP4等磷酸酶催化。虽然人们已经对主要通路的基本过程了解了很多,但是很多详细的信息,如各磷酸化和去磷酸化详细的信号通路和受调控因子等还需要进一步深入研究。已经建立了各种分析检测细胞和组织内的γH2AX的成熟技术和方法,其中免疫发光分析法较为常用。高内涵分析目前是其中较为完善的技术。相比免疫发光分析法,LC/MS是最新发展的一种可绝对定量检测γH2AX的技术,但是其在检测准确性和灵敏度上还需要进一步验证。由于γH2AX可以作为DNA双链断裂、药效、衰老及疾病的潜在分子标志,在体外毒理学和医药学上的研究和应用展示了强大的应用潜力和发展前景,可用于毒物的体外基因毒性高通量初步筛选、疾病诊断、药物药效和疗效评价、药物研发和临床实验研究等。虽然在某些方面还存在一些限制或者问题,但随着相关分子机制更为深入的研究,以及检测分析手段的不断改善,γH2AX在食品、医药、环境、化工、医学等各个领域的发展和应用将会进一步得到更多的提高和推广。
Garcia-Canton C,Anadon A,Meredith C.Toxicol.In Vitro,2012,26(7):1075-1086
Downey M,Durocher D.Cell Cycle,2006,5(13):1376-1381
Rogakou E P,Pilch D R,Orr A H,Ivanova,V S,Bonner W M.J.Biol.Chem.,1998,273(10):5858-5868
West M H P,Bonner W M.Biochemistry,1980,19(14):3238-3245
Fried V A.Science,1992,257(5075):1398-1400
Costanzi C,Pehrson J R.J.Biol.Chem.,2001,276(24):21776-21784
Chadwick B P,Willard H F.J.Cell Biol.,2001,152(2):375-384
Choe J,Kolodrubetz D,Grunstein M.Proc.Nati.Acad.Sci.USA,1982,79(5):1484-1487
Redon C,Pilch D,Rogakou E,Sedelnikova O,Newrock K,Bonner,W.Curr.Opin.Genet.Dev.,2002,12(2):162-169
Tu W Z,Li B,Huang B,Wang Y,Liu X D,Guan H,Zhang S M,Tang Y,Rang W Q,Zhou P K.FEBS letters,2013,587(21):3437-3443
Peak J G,Peak M J.Photochem.Photobiol.,1990,52(2):387-393
Rothkamm K,L brich M.P.Natl.Acad.Sci.USA,2003,100(9):5057-5062
Takahashi A,Mori E,Somakos G I,Ohnishi K,Ohnishi T.Mutat.Res-gen.Tox.En.,2008,656:88-92
Jakob B,Scholz M,&Taucher-Scholz G.Radiat.Res.,2003,159(5):676-684
Hammond E M,Dorie M J,Giaccia A J.J.Biol.Chem.,2003,278(14):12207-12213
Mavrogonatou E,Kletsas D.DNA Repair,2009,8(8):930-943
Xiao H,Li T,Yang J M,Liu L F.Proc.Natl.Acad.Sci.USA,2003,100(9):5205-5210
Tomaso M V D,Basso S,Lafon-Hughes L,Saona G,López-Carro B,Reyes-balos A L,Liddle P.Adv.Biosci.Biotechnol.,2014,5(7):603-616
Garcia-Canton C,Anadon A,Meredith C.Toxicol.Lett.,2013,223:81-87
Sak A,Grehl S,Engelhard M,Wierlemann A,Kaelberlah H P,Erichsen P,P ttgen C,Groneberg M,Stuschke M.Clin.Cancer Res.,2009,15(8):2927-2934
Rybaczek D,Bodys A,Maszewski J.Histochem.Cell Biol.,2007,128(3):227-241
Mischo H E,Hemmerich P,Grosse F,Zhang S.J.Biol.Chem.,2005,280(10):9586-9594
Katsube T,Mori M,Tsuji H,Shiomi T,Wang B,Liu Q,Nenoi M,Onoda M.J.Biochem.,2014:156(2):85-95
Lee Y Y,Yu Y B,Gunawardena H P,Xie L,Chen X. Cell Death Dis.,2012,3(7):1-12
Hunter N,B rner G V,Lichten M,Kleckner N.Nat.Genet.,2001,27(3):236-238
Barral S,Beltramo R,Salio C,Aimar P,Lossi L,Merighi A. Int.J.Mol.Sci.,2014,15(1):1554-1573
Zimmerman E S.Mol.Cell.Biol.,2004,24(21):9286-9294
Daniel R,Ramcharan J,Rogakou E,Taganov K D,Greger J G,Bonner W,Nussenzweig A,Katz R,Skalka A M.J.Biol.Chem.,2004,279(44):45810-45814
Takahashi A,Ohnishi T.Cancer Lett.,2005,229(2):171-179
C'mielová J,Havelek R,Vávrová J,Řezá[AKč]ová,M.Med.Oncol.,2015,32(5):1-8
An J,Huang Y C,Xu Q Z,Zhou L J,Shang Z F,Huang B,Wang Y,Liu X D,Wu D C,Zhou P K.BMC Mol.Biol.,2010,11(18):1-13
Garcia-Cruz R,Roig I,Robles P,Scherthan H,Caldés M G.Reprod.Biomed.Online,2009,18:37-44
Srivastava N,Gochhait S,de Boer P,Bamezai R N K.Mutat.Res.,2009,681(2):180-188
Khoury L,Zalko D,Audebert M.Environ.Mol.Mutagen.,2013,54(9):737-746
Smart D J,Ahmedi K P,Harvey J S,Lynch A M.Mutat.Res-Fund.Mol.M.,2011,715(1):25-31
Banáth J P,Olive P L.Cancer Res.,2003,63(15):4347-4350
Matsuzaki K,Harada A,Takeiri A,Tanaka K,Mishima M.Mutat.Res-gen.Tox.En.,2010,700(1):71-79
FU Li-Wei,CHEN Huan,YANG J,HOU Hong-Wei,HU Qing-Yuan.Chinese J.Anal.Chem., 2014,42(3):320-325付立伟,陈欢,杨进,侯宏卫,胡清源.分析化学,2014,42(3):320-325
Çavaş T,Çinkılıç N,Vatan Ö,Yılmaz D,Coşkun M.Pestic.Biochem.Phys.,2012,104(3):212-217
Ivashkevich A N,Martin O A,Smith A J,Redon C E,Bonner W M,Martin R F,Lobachevsky P N.Mutat.Res-Fund.and Mol.M.,2011,711(1):49-60
Reddig A,Lorenz S,Hiemann R,Guttek K,Hartig R,Heiserich L,Eberle C,Peters V,Schierack P,Sack U,Roggenbuck D,Reinhold D.Cytom.Part A,2015,87(8):724-732
Willitzki A,Lorenz S,Hiemann R,Guttek K,Goihl A,Hartig R,Conrsd K,Feist E,Sack U,Schierack P,Heiserich L,Eberle C,Peters V,Roggenbuck D,Reinhold D.Cytom.Part A,2013,83(11):1017-1026
ZHANG L,DU Guan-Hua.Acta Pharm.Sin.,2005,40(6):486-490 张莉, 杜冠华.药学学报,2005, 40(6):486-490
HU Jing-Yang, YAN Chun-Lin, ZHU Yan, GAO Xiu-Mei. J. Tianjin Univ. Tradit. Chin. Med., 2013,32(2):120-124 胡经阳, 严春琳, 朱彦, 高秀梅.天津中医药大学学报,2013,32(2):120-124
Garcia-Canton C, Anadon A, Meredith C. Mutat. Res-gen. Tox. En., 2013,757(2):158-166
LIU Li-Bo, WANG Li-Li. Chin. J. Pharmacol. Toxicol., 2012,26(6):893-896 刘利波, 王莉莉.中国药理学与毒理学杂志,2012,26(6):893-896
WANG Meng-Meng, HE Ling, HU Mei, ZHAO Chuan. Prog. Pharm. Sci., 2011,35(11):481-486 王萌萌, 何玲, 胡梅, 赵氚.药学进展,2011,35(11):481-486
Matsuda S, Ikura T, Matsuda T. Anal. Bioanal. Chem., 2015:407(18):5521-5527
Sedelnikova O A, Rogakou E P, Panyutin I G, Bonner W M. Radiat. Res., 2002,158(4):486-492
Yu Y, Zhu W, Diao H, Zhou C, Chen F F, Yang J. Toxicol. In Vitro, 2006,20(6):959-965
Zhou C, Li Z, Diao H, Yu Yanke, Zhu Wen, Dai Yayun, Chen F F, Yang J. Mutat. Res-gen. Tox. En., 2006,604(1):8-18
Hanahan D, Weinberg R A. Cell, 2000,100(1):57-70
Kirkland D, Pfuhler S, Tweats D, Aardema M, Corvi R, Darroudi F, Elhajouji A, Glatt H, Hastwell P, Hayashi M, Kasper P, Kirchner S, Lynch A, Marzin D, Maurici D, Meunier J R, Müller L, Nohynek G, Parry J, Parry E, Thybaud V, Tice R, Benthem J V, Vanparrys P, White P. Mutat. Res-gen. Tox. En., 2007,628(1):31-55
L brich M, Shibata A, Beucher A, Fisher A, Ensminger M, Goodarzi A A, Barton O, Jeggo P A. Cell Cycle, 2010,9(4):662-669
Cleaver J E, Feeney L, Revet I. Cell Cycle, 2011,10(19):3223-3224
de Feraudy S, Revet I, Bezrookove V, Feeney L, Cleaver J E. Proc. Natl. Acad. Sci. USA, 2010,107(15):6870-6875
Revet I, Feeney L, Bruguera S, Wilson W, Dong T K, Oh D H, Dankort D, Cleaver J E. Proc. Natl. Acad. Sci. USA, 2011,108(21):8663-8667
Bartkova J, Hořejší Z, Koed K, Krämer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland J M, Lukas C, Ørntoft T, Lukas J, Bartek J. Nature, 2005,434(7035):864-870
CUI Ming, LIU Yang, WANG Guang-Xi, LI Yang, YIN Yu-Xin. Chin. J. Pathol., 2014,43(10):714-717 崔铭, 刘杨, 王光熙, 李扬, 尹玉新.中华病理学杂志,2014,43(10):714-717
Mah L, El-Osta A, Karagiannis T C. Epigenetics, 2010,5(2):129-136
Wang L H, Pfister T D, Parchment R E, Kummar S, Rubinstein L, Evrard Y A, Gutierrez M E, Murgo A J, Tomaszewski J E, Doroshow 70 J H, Kinders R J. Clin. Cancer Res., 2010,16(3):1073-1084
Kinders R J, Hollingshead M, Lawrence S, Ji J, Tabb B, Bonner W M, Pommier Y, Rubinstein L, Evrard Y A, Parcchment R E, Tomaszewski J, Doroshow J H. Clin. Cancer Res., 2010,16(22):5447-5457
Reddig A, Lorenz S, Hiemann R, Guttek K, Hartig R, Heiserich L, Eberle C, Peters Vanessa, Schierack P, Sack U, Roggenbuck D, Reinhold D. Cytom. Part A, 2015,87(8):724-732
Leifert W R, Siddiqui S M. Cytom. Part A, 2015,87(8):692-695

Figure 2 Alignment of amino acid sequences of histone H2A and H2AX for several organisms

Figure 4 Two kinds of processes of dephosphorylation ofγH2AX: (a) dephosphorylation based on the style of histone exchange; (b) dephosphorylation based on kinds of phosphatases

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 下载:
下载:

 下载:
下载:

