
Scheme 1.
Enzymatic production of rubusoside

甜茶苷(Rubusoside, 13-O-β-glucosyl-19-O-β-D-glucosyl-steviol, Ru)是一种稀少的天然甜味剂,是中国广东广西等地特有的野生珍稀蔷薇科悬钩子属植物甜茶的主要甜味成分[1-2],含量约占干叶的5%[2]。 甜茶苷作为甜菊糖苷的一种少量存在于甜叶菊中(少于0.1%)[3]。 目前,尚无化学全合成甜茶苷的报道。 斯替夫苷(Stevioside, 13-O-β-sophorosyl-19-O-β-d-glucosyl-steviol, St)是甜叶菊中两种主要甜菊糖苷之一。 斯替夫苷在分子结构上在其C13位比甜茶苷分子多一个以β-1, 2连接的葡萄糖基。 若能特异性水解斯替夫苷C13上的一个葡萄糖(Scheme 1),即可实现甜茶苷的酶法制备。 但斯替夫苷具有3个以β-糖苷键链接的葡萄糖残基,葡糖基单元的连续水解及重排可能产生甜茶苷、甜菊双糖苷、甜菊醇单葡萄糖酯、甜菊单糖苷、甜菊醇, 异甜菊醇或它们的混合物[4]。
相对于化学合成,酶催化合成可以更专一性地合成目标产物。 本实验室前期以及其它团队已有针对酶促斯替夫苷水解制备甜茶苷的报道[4-8],所使用的酶主要是β-葡萄糖苷酶(EC3.2.1.21)及β-半乳糖苷酶(EC 3.2.1.23)。 Ko等[4-5]使用从Aspergillus aculeatus中获得的β-葡萄糖苷酶催化斯替夫苷水解,甜茶苷的产率为66.07%。 Nguyen等[10]筛选了31种水解酶来水解斯替夫苷制甜茶苷, 其中来自Thermus thermophiles的β-半乳糖苷酶可使斯替夫苷的转化率达到95.4%, 甜茶苷的产率为86%。 Wang等[6]从Streptomyces sp. GXT6分离出一种β-葡萄糖苷酶, 在反应6 h后可使斯替夫苷的转化率达到98.2%, 然而该酶会产生较多的副产物, 甜茶苷的产率为78.8%。 Udompaisarm等[9]通过设计B.thetaiotaomicron VPI-5482的BT_3567β-葡萄糖苷酶基因序列, 表达出一种对斯替夫苷具有高特异性的β-葡萄糖苷酶, 可将甜菊糖苷混合物中的斯替夫苷完全水解。 最近,Lan等[7]报道了一种来自Sphingomonas elodea(ATCC 31461)的β-葡萄糖苷酶, 在反应6 h后斯替夫苷的转化率可达98.6%, 甜茶苷的产率大于99.9%。 我们认为其转化率和收率之间的矛盾可能源于高效液相色谱法(HPLC)的检测偏差。 在他们的实验中,观察到了未表征的物质,在每批反应混合物中该不明物质的HPLC峰面积均增加。 尽管作者认为HPLC谱图中该不明物是原料杂质,但根据该不明物的保留时间及其不断增长的HPLC峰面积,可以猜测该未鉴定的物质可能是Ru继续水解一个葡萄糖基所产生的副产物。 Wan等[8]采用来自Aspergillus sp.的β-半乳糖苷酶水解斯替夫苷, 斯替夫苷的转化率和甜茶苷的产率分别为98.3%和91.4%。 当加酶量增加至5250 U/g时, 反应5 h斯替夫苷的转化率就可达到93.4%, 然而过量的酶导致了更多副产物的出现, 甜茶苷的产率下降到77.58%,该结果也引起了关于反应途径以及过载的酶是否催化副反应的问题。
然而,目前的研究仍存在反应通量不够和副产物不明确等缺点;同时,虽然甜菊糖苷的人体安全性已经得到公认,但对甜茶苷的细胞毒性还鲜有报道。 作为一种甜味剂,甜茶苷摄入人体的主要途径为口服,因此其可能会对人体消化系统如胃、肠、肝等部位造成影响。 许多抗癌药物均具有水溶性差的特点。 药物的糖苷化不仅可以有效增加药物的水溶性,而且可以降低药物的毒性。 已有报道证明甜菊醇具有抑制人体骨肉瘤U2OS细胞增殖的效果[12]。 甜茶苷在C13和C19位比甜菊醇各多出一个葡萄糖基,水溶性比甜菊醇好,也可能具有潜在的抗癌能力但对正常细胞的毒性低。 从安全性或是甜茶苷扩大应用范围这两方面来说,检测甜茶苷对人体细胞,尤其是胃、肠、肝正常细胞与癌细胞的毒性是十分必要的。
本文筛选出来自Aspergillus niger的β-葡萄糖苷酶催化水解斯替夫苷制备甜茶苷,实现了甜茶苷的高通量制备并明确了副产物为甜菊醇单葡萄糖酯。 同时,以传统的化疗药物五氟尿嘧啶为对照,考察了甜茶苷对人消化系统细胞的细胞毒性和甜茶苷的抗癌活性,以便为甜茶苷的应用提供参考。
β-葡萄糖苷酶(ATCC@16404, 来自Aspergillus niger,39000 U/g)由江南大学吴敬教授提供;α-半乳糖苷酶(DS30, 50000 GalU/g)和葡糖淀粉酶(Gluczyme AF6, 1600 U/g)均购自中国天野酶制剂有限公司;淀粉葡萄糖苷酶(A7096, 260 U /mL)和对硝基苯基-β-D-吡喃半乳糖苷(pNPG)均购自中国Sigma-Aldrich公司;斯替夫苷(97%, HPLC)由诸城市浩天药业有限公司提供;胰蛋白酶(生化试剂)、RPMI 1640培养基、DMEM高糖培养基和DMEM/F12培养基均购自美国Gibco公司;胎牛血清购自杭州四季青有限公司;对硝基苯酚(分析纯)、5-氟尿嘧啶(5-FU,生化试剂)、二甲基亚砜(DMSO,分析纯)、碳酸氢钠(分析纯)、氢氧化钠(分析纯)、浓盐酸(分析纯)、乙二胺四乙酸二钠(分析纯)均购自国药集团化学试剂有限公司(上海);聚乙二醇辛基苯基醚(Triton X-100)、核酸内切酶(RNase A)、碘化丙啶(PI,>95%)、噻唑蓝(MTT>98%)、青霉素-链霉素溶液(含100 μg/mL链霉素和100 U/mL青霉素)和Hoechst 33342均购自上海碧云天生物技术有限公司;6孔细胞培养板和96孔细胞培养板均购自美国Costar公司。
人正常肝细胞L02、人盲肠癌细胞HCT-8、人结肠癌细胞HCT 116、人中度分化胃癌细胞SGC-7901、人肝癌细胞BEL-7404和人肝癌细胞BEL-7402均购自中国科学院细胞库;人结肠上皮细胞T84、人胃粘膜上皮细胞GES-1、人肝癌细胞HepG2(P53野生型)、人低分化胃癌细胞MGC-803、人肝癌细胞Huh-7(P53突变型)、人低转移肝癌细胞MHCC97-L、人肝癌细胞Hep3B(P53缺失型)和人高转移肝癌细胞HCCLM3均购自于美国模式培养物集存库 (ATCC)。
2695型高效液相色谱仪 (HPLC,二极管阵列检测器996, 美国Waters公司); Masslynx色谱工作站;Avance Ⅲ 600型核磁共振谱仪(NMR,德国Bruker公司);FACSAria型流式细胞仪(美国Becton Dickinson公司);3111型二氧化碳培养箱(美国Thermo Epectron Corporation公司);XDS-1A型倒置显微镜(上海精密仪器仪表有限公司);H4-200型荧光显微镜(日本Olympus 光学仪器公司);离心机(RJ-TDL-40B,无锡市瑞江分析仪器有限公司);μQuant型酶标仪(美国Bio-Tek公司);液相色谱串联四极杆飞行时间质谱仪(LC-MS,美国Waters公司);Lambda 25型紫外可见分光光度计(UV-Vis,美国Perkin Elmer公司)。
酶的筛选 在1%的斯替夫苷水溶液中加入不同的糖苷酶,分别在各种酶对应的最适温度下振荡(150 r/min)反应24 h,然后用与反应液等体积的甲醇终止反应。 用HPLC分析反应液。 HPLC检测条件如下:APS-2 HYPERSIL 氨基色谱柱,柱温30 ℃;梯度洗脱条件如下:80%乙腈(0~2 min), 80%~50%乙腈(2~16 min), 50%~80%乙腈(16~20 min), 80%乙腈(20~25 min)。 用LC-MS分析反应液中各组分的相对分子质量。
β-葡萄糖苷酶酶活测定 用pNPG作为底物测定β-葡糖苷酶活性。 将100 μL 1 mg/mL酶液加入800 μL柠檬酸-磷酸氢二钠缓冲液(pH=6, 10 mmol/L)中。 60 ℃下预热上述混合液10 min后, 加入100 μL pNPG溶液(5 mmol/L)开始反应, 反应10 min后用2 mL 1 mol/L Na2CO3终止反应。 使用UV-Vis测量反应液在405 nm处的吸光度,根据对硝基苯酚(pNP)标准曲线计算反应生成pNP的量。 将不含酶的反应混合物用作空白对照。 一个单位的β-葡糖苷酶活性(U)定义为在60 ℃下,每 min水解pNPG生成1 nmol对硝基苯酚所需的酶量。
将斯替夫苷溶液(100 g/L)在60 ℃水浴保温30 min;然后加入β-葡萄糖苷酶(300 U/g斯替夫苷), 150 r/min下震荡反应24 h。 定时取样, 用同等体积的甲醇终止反应。 斯替夫苷的转化率(C(St))和甜菜苷的产率(Y(Ru))计算如下:
|
$ C\left( {{\rm{St}}} \right){\rm{ = }}\left( {{\rho _0} - {\rho _t}} \right)/{\rho _0} \times 100\% $ |
(1) |
|
$ Y\left( {{\rm{Ru}}} \right){\rm{ = }}\frac{{{\rho _{{\rm{RU}}}}/{M_{{\rm{RU}}}}}}{{{\rho _0}/{M_{{\rm{St}}}}}} \times 100{\rm{\% }} $ |
(2) |
式中,ρ0(g/L)是反应液中St的初始质量浓度, ρt(g/L)是t时刻反应液中St的质量浓度,ρRu(g/L)是反应液中Ru的质量浓度,MRu=642,MSt=804。 用HPLC检测反应液中的Ru和St的质量浓度。 根据其色谱面积百分比确定并计算每种产物的产率, 并用斯替夫苷的标准曲线校准。 分别以50%甲醇和95%甲醇为重结晶溶剂,分步纯化产物。
将细胞接种于含100 U/mL青霉素、100 mg/mL链霉素和10%的胎牛血清的培养基中,置于37 ℃、5%CO2培养箱中培养。 根据不同细胞生长需要选用不同的培养基,其中SGC-7901细胞用RPMI 1640培养基,T84细胞用DMEM/F12培养基,其余细胞用DMEM高糖培养基。
将细胞于37 ℃、5%CO2培养箱中培养至对数生长期。 用0.25%的胰蛋白酶消化对数生长期细胞。 当细胞浓度达到5×104个/mL时,接种细胞于96孔板中,每孔接种量为100 μL。 加入150 μL溶有甜茶苷的培养基,对照组以新培养基或者含有0.5%DMSO的新培养基替代甜茶苷溶液,设6个复孔作为平行样。 加药培养48 h后,每孔加入100 μL 0.5 mg/mL MTT溶液,在37 ℃下培养4 h后吸弃上清液,每孔加入150 μL DMSO,在微量振荡器上振荡10 min,用酶标仪在570 nm下测定每孔所对应的吸光度,通过式(3)计算细胞存活率(S%)。
|
$ S/\% = \left( {{A_{{\rm{Experiment}}}}/{A_{{\rm{Control}}}}} \right) \times 100{\rm{\% }} $ |
(3) |
式中,AExperiment为经药物作用后的吸光度值,AControl为未经药物作用的吸光度。
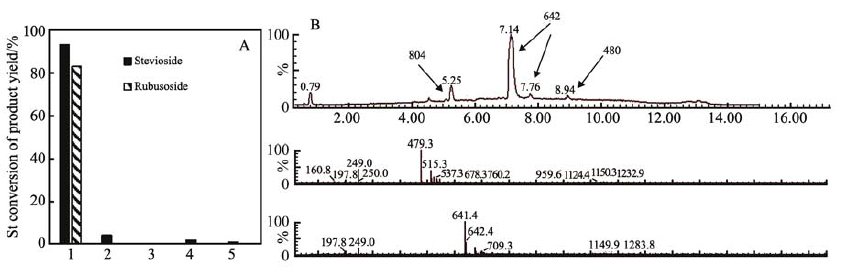
5种水解酶催化斯替夫苷的活性如图 1A所示。 只有来自Aspergillus niger的β-葡萄糖苷酶能够水解St且产物较为单一。 通过LC-MS对水解产物进行进一步表征(图 1B),主产物的相对分子质量为642,副产物的相对分子质量为480,通过LC-MS及核磁表征确定主要的副产物为甜菊单葡萄糖酯(分析数据见辅助材料图S1,S2,S3)。
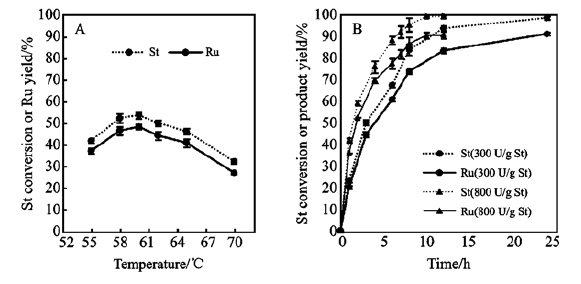
温度、反应时间以及加酶量对斯替夫苷水解的影响如图 2所示,该酶最佳的水解温度为60 ℃,在该温度下,当加酶量为0.8%(300 U/g St)时,24 h下甜茶苷的产率为90.8%,斯替夫苷的转化率达98.2%;当加酶量为2%(800 U/g St)时,12 h下甜茶苷的产率为90.4%,斯替夫苷的转化率达98.8%。 该反应的副产物较少,方便后续对甜茶苷的分离与提纯,其次所需加酶量较小。
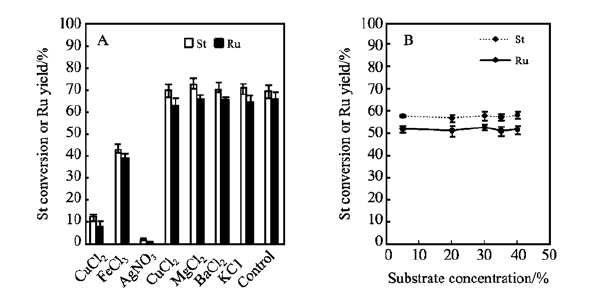
金属离子对于酶的稳定性以及酶活具有重要影响。 如图 3A所示,与未添加金属离子的空白组对比发现,5 mmol/L的Mg2+、Ca2+、K+对斯替夫苷的水解具有一定的促进作用,而同浓度下的Fe3+、Ag+、Cu2+对β-葡萄糖苷酶具有明显的抑制作用,在实际生产中应尽量避免使用此类材质的反应釜,例如品质较差的不锈钢釜。 图 3B证明该反应不存在底物抑制,该特性有利于工业上甜茶苷的大规模生产。
分别用50%甲醇, 95%甲醇分两步重结晶,得到HPLC纯度大于99%的主水解产物,其具有与天然甜茶苷相同的保留时间及相对分子质量。 然而,甜菊糖苷属化合物存在一些同分异构体,它们的保留时间也可能相同。 因此,仅通过LC-MS以及HPLC无法准确判断水解产物的结构。
主水解产物的一维核磁共振谱如图 3所示,化学位移范围为3~5.5的峰来自糖单元, 化学位移小于2.3的峰来自糖苷配基。 图 3B中有4个峰值, 包括5.39(d, J=8.1 Hz, 1H), 5.15(s, J=1.5 Hz, 1H), 4.85(s, 1H), 4.51(d, J=7.8 Hz, 1H), 只有两个峰出现双重峰分裂, 高J耦合距离表明这两个糖基为β构型, 5.15和4.85的峰未显示出分裂, 是糖苷配基的信号峰。 1.21和1.01的峰有相对高的强度, 归属糖苷配基的18和20位上—CH3的质子。 糖苷配基团的质子化学位移标记在图 3C中。 图 3D为化合物的13C谱。 糖单元的化学位移主要分布在60~103的范围内, 而糖苷配基的光谱大多位于10~60的范围内。 由于羧基(C19位)和C=C双键(16和17位), 在光谱中的低场也观察到3个峰(19, 177.1;16, 153.4和17, 103.4)。 核磁共振谱的特征峰与文献报道[11]的甜茶苷的特征峰一致。 结合LC-MS结果以及保留时间, 可以确定该产物是甜茶苷,分子结构见Scheme 1。
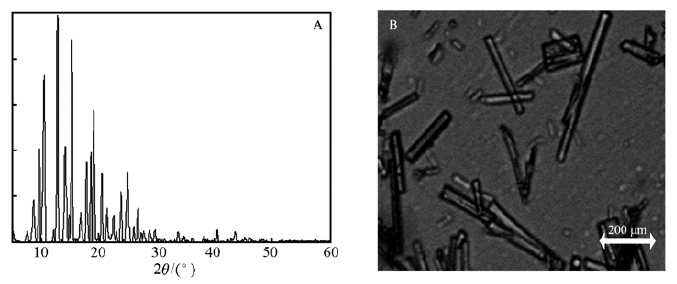
如图 4A为纯度95%的甜茶苷晶体。 在2θ=8.7°、9.7°、10.5°、12.8°、14.1°、14.8°、15.3°、16.9°、17.8°、18.6°、19.1°、20.5°、21.4°、22.5°、23.9°、24.9°和26.7°出现了衍射峰。 采用Jade6.5软件分析XRD数据,采用多峰分离法计算甜茶苷晶体的结晶化度,计算得各峰的结晶化度均高于非晶体的判断阈值,表明甜茶苷的结晶度高。 图 4B为甜茶苷晶体的形貌,甜茶苷晶体呈棒状结构,表面均匀,结晶化程度高。
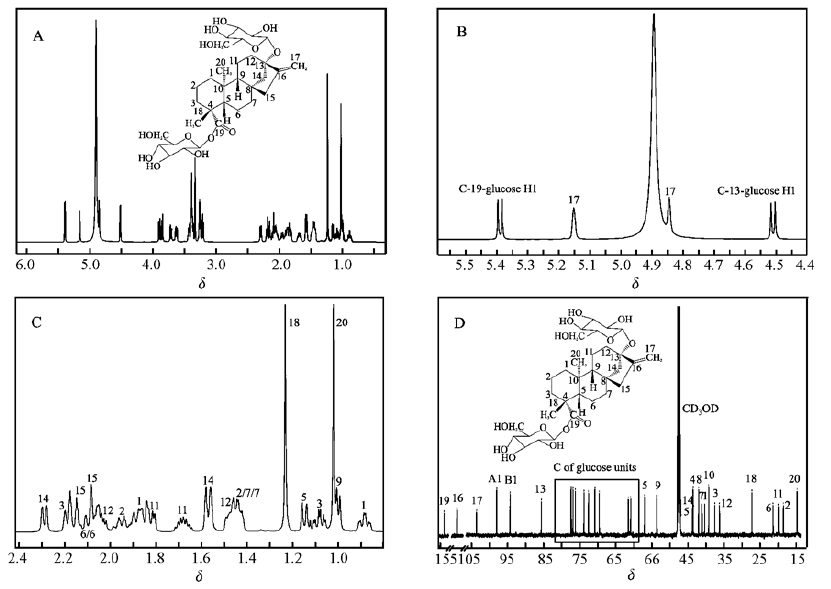
A.1H NMR spectrum; B.1H Enlarged anomeric region for sugar units; C.1H enlarged proton NMR for aglycone part; D.13C NMR spectrum
如图 5所示,甜茶苷作用下的人体正常胃(CES-1)、肠(T84)、肝(L02)细胞的存活率均高于90%,表明甜茶苷对于正常细胞的细胞毒性为零级,即无细胞毒性。 250 μg/mL的甜茶苷对于人肝癌细胞BEL-7404的抑制效果较明显,为同质量浓度下5-氟尿嘧啶(5-Fu)效果的30%。 250 μg/mL的甜茶苷对癌细胞无太强的抑制作用,在细胞毒性评级上同属零级,然而其作为一种天然安全的甜味剂,与典型化疗药物5-氟尿嘧啶相比具有更低的毒性以及更好的溶解度,增加使用的剂量不会造成毒副影响,在高质量浓度下具有一定抑制癌细胞生长的潜力。
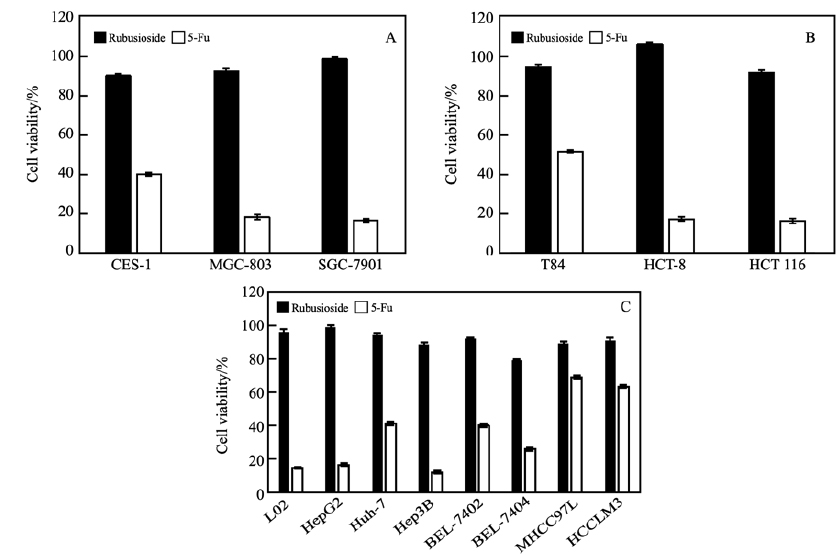
A.stomach cells; B.intestinal cells; C.liver cells
通过核磁解析, 确定了来自Aspergillus niger的β-葡萄糖苷酶能特异性水解斯替夫苷C13位上的一个葡萄糖从而生成甜茶苷。 当加酶量为300 U/g斯替夫苷时, 60 ℃下反应24 h, 斯替夫苷转化率可达98.2%, 甜茶苷的产率为90.8%;当加酶量增大到800 U/g斯替夫苷时, 60 ℃下, 反应10 h转化率即可达98.8%, 产率为90.1%,反应产物单一, 副反应少。 采用甲醇水溶液通过重结晶操作可获得纯度大于95%的甜茶苷晶体,且晶体的结晶度高。 此外,甜茶苷无细胞毒性,是一种安全的甜味剂,250 μg/mL的甜茶苷对人体肝癌细胞BEL-7404有一定的抑制效果,抑制力为同质量浓度下5-氟尿嘧啶的30%。
辅助材料(Supporting Information)[副产物甜菊醇单葡萄糖酯的核磁共振谱图]可以免费从本刊网站(http://yyhx.ciac.jl.cn/)下载。
Ohtani K, Aikawa Y, Kasai R. Minor Diterpene Glycosides from Sweet Leaves of Rubus suavissimus[J]. Phytochemistry, 1992, 31(5): 1553-1559.
Koh G Y, Chou G, Liu Z. Purification of a Water Extract of Chinese Sweet Tea Plant (Rubus suavissimus, S. Lee) by Alcohol Precipitation[J]. J Agric Food Chem, 2009, 57(11): 5000-5006.
Uhler B, Yang Z. Rebaudioside A and Other Unreported Steviol Glycoside Isomers Found in the Sweet Tea (Rubus suavissimis) Leaf[J]. Phytochem Lett, 2018, 28: 93-97.
Ko J A, Ryu Y B, Kwon H J. Characterization of a Novel Steviol-Producing β-Glucosidase from Penicillium Decumbens and Optimal Production of the Steviol[J]. Appl Microbiol Biotechol, 2013, 97(18): 8151-8161.
Ko J A, Kim Y M, Ryu Y B. Mass Production of Rubusoside Using a Novel Stevioside-Specific beta-Glucosidase from Aspergillus aculeatus[J]. J Agric Food Chem, 2012, 60(24): 6210-6216.
Wang Z L, Wang J P, Jiang M H. Selective Production of Rubusoside from Stevioside by Using the Sophorose Activity of beta-Glucosidase from Streptomyces sp GXT6[J]. Appl Microbiol Biotechnol, 2015, 99(22): 9663-9674.
Lan Q, Tang T T, Yin Y. Highly Specific Sophorose beta-Glucosidase from Sphingomonas elodea ATCC 31461 for the Efficient Conversion of Stevioside to Rubusoside[J]. Food Chem, 2019, 295: 563-568.
Wan H D, Tao G J, Kim D. Enzymatic Preparation of a Natural Sweetener Rubusoside from Specific Hydrolysis of Stevioside with beta-Galactosidase from Aspergillus sp.[J]. J Mol Catal B-Enzym, 2012, 82: 12-17.
Udompaisarn S, Arthan D, Somana J. Development and Validation of an Enzymatic Method to Determine Stevioside Content from Stevia rebaudiana[J]. J Agric Food Chem, 2017, 65(15): 3223-3229.
Nguyen T T H, Jung S J, Kang H K. Production of Rubusoside from Stevioside by Using a Thermostable Lactase from Thermus thermophilus and Solubility Enhancement of Liquiritin and Teniposide[J]. Enzyme Microb Technol, 2014, 64/65(7): 38-43.
Sugimoto N, Sato K, Liu H M. Analysis of Rubusoside and Related Compounds in Tenryocha Extract Sweetener[J]. J Food Hyg Soc Jpn, 2002, 43(4): 250-253.
Chen J M, Zhang J, Xia Y M. The Natural Sweetener Metabolite Steviol Inhibits the Proliferation of Human Osteosarcoma U2OS Cell Line[J]. Oncol Lett, 2018, 15(4): 5250-5256.
Chen J M, Ding L, Sui X C. Production of a Bioactive Sweetener Steviolbioside via Specific Hydrolyzing Ester Linkage of Stevioside with a beta-Galactosidase[J]. Food Chem, 2016, 196: 155-160.

图 1 (A)酶促水解斯替夫苷。 10 mg/mL,12 h。 1.来自Aspergillus niger的β-葡萄糖苷酶, 60 ℃, 1%(质量分数), 去离子水;2.β-半乳糖苷酶(Lactozym Pure 7500 L), 37 ℃, 12%(质量分数), 去离子水; 3.α-半乳糖苷酶(DS30), 60 ℃, pH=5.0, 2%(质量分数); 4.淀粉葡萄糖苷酶(A7096), 60 ℃, 1%(质量分数), 去离子水; 5.葡糖淀粉酶(Gluczyme AF6), 60 ℃, 2%(质量分数), 去离子水; (B)水解产物的LC-MS图
Figure 1 (A)Enzymatic hydrolysis of stevioside at 10 mg/mL in 12 h. 1.β-glucosidase from Aspergillus niger, 60 ℃, 1%, DI water; 2.β-galactosidase (Lactozym Pure 7500L), 37 ℃, 12%, DI water; 3.α-galactosidase (DS30), 60 ℃, 2%, pH=5.0; 4.Amyl glucosidase (A7096), 55 ℃, 1%, DI water; 5.glucoamylase (Gluczyme AF6), 2%, 60 ℃, DI water; (B)LC-MS spectrum of the hydrolysis product

图 2 (A)反应温度对斯替夫苷水解的影响(0.2 g St/mL, 300 U/g St, 3.5 h);(B)加酶量与反应时间对斯替夫苷水解的影响( 0.2 g St/mL, 60 ℃)
Figure 2 (A)Temperature effect (0.2 g St/mL, 300 U/g St, 3.5 h); (B)Time course of the hydrolysis (0.2 g St/mL, 60 ℃)

图 3 (A)金属离子对斯替夫苷水解的影响(0.01 g St/mL, 300 U/g St, 5 mmol/L金属离子, 6 h);(B)底物浓度对斯替夫苷水解的影响(300 U/g St, 60 ℃, 5 h)
Figure 3 (A)Electrolyte effect(0.01 g St/mL, 300 U/g St, 5 mmol/L metal ions, 6 h); (B)Substrate concentration effect(300 U/g St, 60℃, 5 h)

图 4 甜茶苷的一维NMR分析
Figure 4 1H NMR analysis of rubusoside
A.1H NMR spectrum; B.1H Enlarged anomeric region for sugar units; C.1H enlarged proton NMR for aglycone part; D.13C NMR spectrum

图 5 甜茶苷晶体的XRD谱图(A)和)光学图片(B)
Figure 5 (A)XRD spectrum and (B)optical image of the rubusoside crystal

 扫一扫看文章
扫一扫看文章

扫一扫关注我们
 下载:
下载:

 下载:
下载:
