Ningbo University's Faculty of Science, Ningbo University, Ningbo 315211, China;
2.
School of Life Science, Inner Mongolia Agricultural University, Hohhot 010018, China;
3.
School of Physics and Information Technology, Shaanxi Normal University, Xi'an 710062, China;
4.
Division of Physical Biology & Bioimaging Center, Shanghai Synchrotron Radiation Facility, Chinese Academy of Sciences, Key Laboratory of Interfacial Physics and Technology, Shanghai Institute of Applied Physics, Chinese Academy of Sciences, Shanghai 201800, China
Received Date:
09 February 2016 Revised Date:
12 May 2016
Abstract:
Atomic force microscopy (AFM) is one of important tools for studying the structure and function of biomolecules due to its nanoscale capability. A well prepared sample was the primary key issue for high-resolution imaging macromolecules. Here, by using self-assembly technology of DNA origami, antigens were modified on DNA origami. Afterward, antibodies reacted specifically with antigens by a molecular recognition way. Then, a nanostructure formed from DNA origami and antigen-antibody complexes. By the aid of adsorption character of DNA origami, individual antibodies could selectively adsorb on mica surface. As a result, the ultrastructural morphology image of a single IgG molecule, the characteristic "Y-shaped" domains, was resolved in liquid. The way for adsorbing biomolecules on the mica surface in a liquid environment was simple and convenient, and it may be useful to detect and measure biomoleculers at the singe-molecule level with AFM.
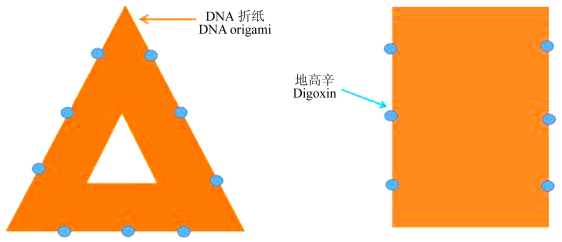
Figure 1.
Schematic presentation of positions of digoxin molecules modified on DNA origami nanostructures. There were 6 digoxin molecules fixed on one side of DNA origami. Two digoxin molecules formed a couple binding site within a distance of 8-18 nm. Left: positions of digoxin molecules on the triangular-shaped DNA origami; Right: positions of digoxin molecules on the rectangular-shaped DNA origami
Figure 3.
Images of individual IgG molecules. (a) AFM image of single IgG molecule binding to the triangular-shaped DNA origami; (b) enlarged AFM image in (a) marked with white square. The top: AFM image of single IgG molecule; the bottom, the project image of the IgG in (b) marked in yellow.
Figure 4.
Images of individual IgG molecules. (a) AFM image of single IgG molecule binding to the rectangular-shaped DNA origami; (b) enlarged AFM image in (a) marked with white square. The up: AFM image of single IgG molecule; The bottom left and middle, the projected images of IgG in (b) marked in red and blue; The bottom right, the projected image (yellow) of IgG in (b) of Fig 3.
KrissanaprasitA,Madsen M,Knudsen J B,Gudnason D,Surareungchai W,Birkedal V,Gothelf K V.ACS Nano,2016,10(2):2243-2250
[21]
Ding B Q,Deng Z D,Yan H,Cabrini S,Zuckermann R N,Bokor J. J.Am.Chem.Soc.,2010,132(10):3248-3249
[22]
Ke Y G,Lindsay S,Chang Y,Liu Y,Yan H.Science,2008,319(5860):180-183
[23]
Wu N,Czajkowsky D M,Zhang J J,Qu J X,Ye M,Zeng D D,Zhou X F,Hu J,Shao Z F,Li B,Fan C H.J.Am.Chem.Soc.,2013,135(33):12172-12175
Figure 1
Schematic presentation of positions of digoxin molecules modified on DNA origami nanostructures. There were 6 digoxin molecules fixed on one side of DNA origami. Two digoxin molecules formed a couple binding site within a distance of 8-18 nm. Left: positions of digoxin molecules on the triangular-shaped DNA origami; Right: positions of digoxin molecules on the rectangular-shaped DNA origami
Figure 3
Images of individual IgG molecules. (a) AFM image of single IgG molecule binding to the triangular-shaped DNA origami; (b) enlarged AFM image in (a) marked with white square. The top: AFM image of single IgG molecule; the bottom, the project image of the IgG in (b) marked in yellow.
Figure 4
Images of individual IgG molecules. (a) AFM image of single IgG molecule binding to the rectangular-shaped DNA origami; (b) enlarged AFM image in (a) marked with white square. The up: AFM image of single IgG molecule; The bottom left and middle, the projected images of IgG in (b) marked in red and blue; The bottom right, the projected image (yellow) of IgG in (b) of Fig 3.

 下载:
下载:




 下载:
下载:





